Труды государственногозаповедника "Столбы". Выпуск III. Скрытностволовые вредители леса на гарях государственного заповедника «Столбы»
Введение
Исследования по данному вопросу проводились в 1949 и 1950 годах и до настоящего времени являются единственной работой по энтомологии в лесах заповедника «Столбы». Естественно-историческая и климатическая характеристика этих лесов дана во II томе «Трудов заповедника».
Территория заповедника расположена в южной части Средней Сибири в северо-западных отрогах Восточного Саяна и отличается своеобразием энтомофауны, еще недостаточно исследованной.
Для изучения нами были взяты гари в районе Столбинского нагорья в кварталах 31 и 48, в наиболее распространенных типах горно-таежных лесов, а именно — сосняках-черничниках и пихтарниках-зеленомошниках. Эти гари имеют 7,15 и 23-летнюю давность. Более молодых гарей не имеется.
В дополнение к этому району нами исследовались сосновые и еловые гари 1945 года в районе р.Слизневой, а также гарь 1948 года в сосняке разнотравном предгорной части заповедника в районе ручья Лалетина, недалеко от северо-западной границы заповедника.
Изучались вредители хвойных пород из семейства Jpidae, Cerambycidae, Curculionidae (род Hylobius u Pissodes), Siricidae и отчасти Buprestidae.
Исследовался видовой состав этих вредителей (по связи вертикальной зональности), биоэкологические особенности ведущих видов в данном географическом районе и их динамика по годам. Изучались биология и влияние на численность вредителей насекомых, являющихся их естественными врагами.
Предполагалось также наметить мероприятия по борьбе с вредителями и предупреждению их массового размножения, применимые в условиях заповедного режима.
Метод исследования. Наблюдения проводились на постоянных пробных площадях в насаждениях, пройденных пожаром, а также не горевших. Последние взяты условно, так как за исключением приручейных типов леса, все насаждения заповедника в более или менее отдаленное время горели.
В горно-таежном поясе (район Столбинского нагорья) были заложены 9 постоянных проб; в переходном поясе к предгорьям, в районе реки Слизневой и ручья Лалетина, 2 постоянные пробы и 2 временные. Всего было взято 13 проб, в том числе 7 на сосновых гарях, 2 на пихтовых, 2 на еловых гарях и по одной контрольной пробе в сосняке-черничнике и пихтарнике-зеленомошнике — наиболее распространенных типах леса.
На каждой пробной площади в 1949 и повторно в 1950 годах согласно методике А.В.Яцентковского (1931) произведено энтомологическое описание зараженных деревьев по годам усыхания, а также сухостоя, имевшегося до пожара. Поскольку точное определение времени усыхания деревьев затруднительно, пришлось весь сухостой после пожара разделить на две группы: средний и старый. К среднему сухостою относились условно все деревья, усохшие за 3 года, предшествовавшие 1948 году, к старому весь остальной сухостой, накопившийся с момента пожара. Каждой группе сухостоя присущи свои характерные особенности состояния коры и кроны по степени сохранности и свежести. Как оказалось, старый сухостой большей частью слабо заражен вредителями вследствие небольшого первоначального запаса вредителей на гарях. Средний сухостой, как правило, густо заселен вредителями, так как короеды и усачи особенно сильно размножаются на третий, четвертый и пятый год после пожара. Бурелом и ветровал в каждой группе учитывался особо.
Степень зараженности и видовой состав вредителей на гарях исследовались на модельных деревьях, по возможности на полный анализ, в зависимости от степени сохранности коры.
Модельные деревья, взятые из сухостоя и повала, количественный анализ зараженности которых был неосуществим, использовались только для изучения видового состава, последовательности заселения, стаций поселения. На срубленных свежезараженных деревьях, путем взятия палеток коры, велись систематические исследования биологии вредителей, наблюдения над последовательностью заселения вредителями, фазами их развития и над деятельностью их врагов — насекомых и птиц.
Всего было взято 104 модельных дерева сосны, ели, пихты, лиственницы и кедра. Из них 38 стоящих деревьев, в том числе зараженные, усыхающие и усохшие и 66 поваленных. В числе последних 21 дерево было срублено в качестве ловчих в 1950 году, а 45 из естественного ветровала и срубленных осенью 1948 года при устройстве квартальной сети.
На полный анализ было взято 34 модели. Анализ производился по двухметровым отрубкам ствола отдельно по секторам: стоящих зараженных — по странам света, поваленных — по верхнему, нижнему и боковым секторам с отметкой степени их затененности в зависимости от положения ствола. На каждой модели палетки коры брались с учетом видового состава — величиной от 1 кв.дм до размеров во всю ширину сектора и протяжением до 0,5 м. Анализировались также ветви и частично корни.
Вредная деятельность лубоедов рода Blastofagus и усачей рода Monohamus по подстрижке побегов сосны на пробных площадях и вне их, в сосняках-черничниках и сосняках-брусничниках исследовалась в 1949-1950 годах на 32 учетных площадках размером по 25 кв.м. На пробных площадях исследовалось также естественное возобновление сосны и других хвойных для учета вредной деятельности долгоносиков рода Hylobius u Pissodes.
В 1950 году широко применялся метод садков — сетчатых, глухих деревянных с закладкой метровых отрубков зараженных стволов, а также матерчатых садков, стеклянных банок и проч. с закладкой частей тонких стволов и ветвей для вывода насекомых. Каждый вид насекомых собирался в достаточном количестве для коллекционирования и определения.
Видовой состав насекомых вредителей и их паразитов и хищников определялся автором (см. таблицы видового состава 1 и 2) с последующими консультациями у специалистов-систематиков. В частности, единственный вид из рода Saperda, выведенный на ели, а именно S. interrupta Gebl. — определен Н.Н.Плавильщиковым.
Автор глубоко признателен В.Н.Степанову за оказанную помощь советами и за контрольное определение сборов из сем. Buprestidae и некоторых других семейств.
1. Зоогеографическая характеристика скрытностволовых вредителей на гарях заповедника
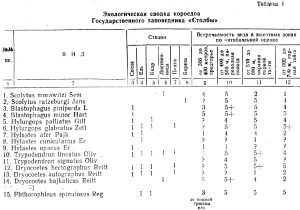
Обзор видового состава ипидофауны (таблица 1) позволяет выделить среди 36 видов следующие зоогеографические элементы:
1. Видов широкого евразийского распространения насчитывается 21 (58 проц.).
2. Видов, близких к первой группе, но с более отодвинутой к северу границей распространения — 12 (34 проц.).
3. Видов сибирских, но частично заходящих в северные районы Европейской части Союза — 3 (8 проц.).
В списке ипидофауны можно выделить 16 видов, наиболее многочисленных и широко распространенных в заповеднике. В их числе 7 видов (44 проц.) из первой группы, 8 видов (50 проц.) из второй группы и 1 вид (6 проц.) из третьей.
В числе массовых видов короедов, изучение которых имеет наибольшее хозяйственное значение, находятся Blastofagus piniperda, В. minor u Pityogenes chalcographus относятся к первой группе, a Jps typographus, Polygraphus subopacus, Xylechinus pilosus — ко второй.
Отражением довольно суровых климатических условий заповедника является относительно высокий процент северных палео-бореальных видов.
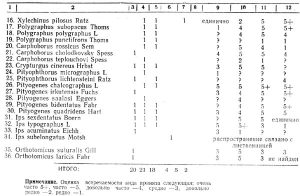
Видовой состав сем. Cerambycidae (таблица 2) в основном также относится к палеарктической фауне. По зоогеографическим элементам его можно охарактеризовать следующим образом (таблица 3).
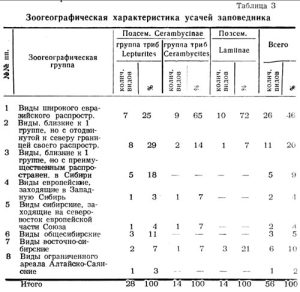
В целом по семейству европейско-азиатские виды составляют 75 проц., европейские — 4 проц., сибирские — 21 проц.
По отдельным систематическим группировкам это соотношение не выдерживается. Наиболее высоко стоящее в своем развитии подсемейство Lamiinae представлено 79 проц. видов широкого европейско-сибирского происхождения, в том числе всего 7% северных видов. В подсемействе Cerambycinae, группе триб Сеrambycites процент северных палеобореальных выше вдвое, а в группе триб Lepturites выше вчетверо, чем в подсемействе Lamiinae. Из таблицы 1 видно, что Lepturites включает в себя все 8 выделенных нами зоогеографических групп, тогда как Cerambycites только 5, а подсемейство Lamiinae — 3.
Следовательно, в лесах заповедника, чем совершеннее отдельные систематические группы усачей в своем развитии, тем выше среди них процент видов широкого евразийского распространения, тем меньше процент видов близких к ним, но дальше заходящих к северу, и одновременно среди них исчезают виды суженных ареалов и эндемиков.
Для групп триб Lepturites характерно наличие 5 видов, которые обычны в Сибири, но редки или встречаются спорадически в европейской части Союза (3 группа), и которые с типично азиатскими видами составляют 12 видов или 43 проц.
Типичными сибирскими видами являются Evodinus variabilis Gebl, Acmaeops angusticollis Gebl, Leptura sequensi Reitt, Stran­galia duodecimguttata F. более многочисленная, по мере при­ближения к востоку, Chlorophorus gracilipes Fald, Clutus arietoides Reitt, Strangalia nigripes rufipennis Bless — восточно-сибирский подвид, встречающийся в заповеднике совместно с европейским подвидом Strangalia nigripes Deg.
Часто встречающиеся в Восточной Сибири восточно-сибирские а также сибирские виды: Allosterna tabacicolor bivitis Motsch, Tetropium gracilicorne Reitt, Monochamus impluviatus Motsch, Acanthocinus carinulatus Gebl.
Несомненно, что вследствие редкой здесь встречаемости часть восточно-сибирских, а также европейских видов могла выпасть из наших сборов.
Из других исследованных семейств вредителей представители восточно-сибирской энтомофауны встречены только среди рогохвостов, а именно — Sirex umbra Sem, свойственный Амурскому и Уссурийскому краям и Paururus ermak Sem, характерный для Забайкалья, Дальнего Востока, Сахалина.
На основании зоогеографического анализа сем. Cerambycidae в заповеднике «Столбы» можно сделать следующее заключение.
На фоне видов широкого европейского распространения количество видов, заходящих с востока, и частота их встречаемости значительно выше, чем заходящих с запада. Следовательно, заповедник, расположенный в южной части Средней Сибири и отграниченный от Западной Сибири всего лишь рекой Енисей, по своему естественному природному комплексу ближе к Восточной Сибири. С другой стороны, хорошо представленное количество северных видов позволяет судить о довольно суровых природных условиях данной местности.
2. Биоэкологический очерк скрытностволовых вредителей на гарях заповедника
Видовой состав и численность скрытностволовых вредителей в обследованной нами части заповедника зависит главным образом от следующих факторов:
1. От территориального распределения основных древесных пород, как стации поселения вредителей, от преобладания спелых и приспевающих насаждений, отсутствия свежих вырубок в силу особенностей заповедного режима.
2. От типа леса и изменений, вносимых в природную обстановку бывшими пожарами и прежними рубками.
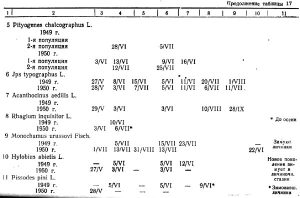
3. От разнообразия экологических условий, создаваемых горным рельефом; распределения тепла и влаги, чрезвычайно меняющейся вследствие этого на очень небольшом пространстве растительности. Большое значение имеет высота над уровнем моря, экспозиция склонов, близость ручьев с их долинами, узкими и слабо разработанными в горно-таежном районе и более широкими с крутыми склонами в предгорьях. Разнообразие экологических условий в горах приводит к неравномерному созреванию imago отдельных видов вредителей, растянутости сроков лета. У некоторых видов, как например Polygraphus polygraphus L u Pityogenes chalcographus L, наблюдаются две фенологические популяции.
Несмотря на сравнительно небольшую разницу по высоте над уровнем моря в крайних точках исследованного района — всего 400 м — влияние вертикальной зональности весьма ощутимо. С поднятием на каждые 100 м, наряду с изменением климатических условий, изменяется и количественное распределение отдельных групп насекомых на гарях и вне их. В качестве примера приводим экологическую характеристику ипидофауны (таблица 1).
Ведущие виды вредителей сосны (Pinus silvestris L) на гарях горно-таежного района — Blastophagus piniperda L, В. minor Hart, Monochamus urussovi Fisch, Hylobius abietis L и Pissodes pini L обычны во всех экологических условиях. Однако каждому виду присущи свои биологические и экологические особенности применительно к данным условиям внешней среды.
В. minor — малый садовник особенно распространен в сосняках переходного пояса между Столбинским нагорьем и примыкающими предгорьями в пределах высот от 400 до 600 м над уровнем моря, а также в предгорных сосняках разнотравных, особенно вблизи реки Слизневой и ручья Лалетина и впадающих в них ручьев. Наибольшие его концентрации наблюдаются на гарях до 15-летней давности.
Массовый лет его в заповеднике по двухлетним наблюдениям начинается с середины мая; появление личинок в первых числах июня; окукливание в третьей декаде июля. Молодые жуки отмечены в конце июля — начале августа. Вылет молодых жуков, связанный с достаточным отвердением хитиновых покровов, в некоторые годы может задерживаться из-за плохой погоды; обычно же происходит в августе. К концу августа под корой остаются единичные запоздавшие жуки.
В 1949 году, отличавшемся дождливой и холодной погодой, окукливание наблюдалось везде в толще коры, за исключением верхней части ствола с тонкой корой, где куколочные колыбельки располагались на глубине 1-3 мм. В 1950 году личинки окукливались, как правило, в древесине, на глубине 7-8 мм и только при достаточно толстой коре окукливались в последней.
Самое ранее формирование жуков (наблюдалось на солнечной стороне при толщине коры от 3 до 4 мм. Но в 1950 году в этих же условиях на некоторых хорошо освещенных моделях наблюдалась сплошная гибель личинок вследствие перегрева и пересыхания верхних слоев древесины.
Под толстой корой и особенно с нижней стороны лежащего ствола вылет молодых жуков задерживался на 2-3 недели по сравнению с общей массой жуков. В 1949 году единичные личинки и куколки наблюдались в толстой коре 9-12 сентября, а в 1950 году — даже в конце сентября.
В конце августа в приспевающих и спелых сосняках начинают опадать первые подстриженные садовником побеги и продолжают опадать до конца октября и позже. Молодые жуки обнаруживаются в побегах даже в это позднее время. Кроме сосны, садовник изредка использует для дополнительного питания пихту, ель, лиственницу, кедр.
Малый садовник охотно заселяет свежеповаленные деревья в изреженных, хорошо освещенных сосняках. В 1949 году шесть моделей сосны, срубленные осенью 1948 года при устройстве квартальной сети и взятые на визире подряд, оказались заселенными малым садовником, из них пять моделей в сильной степени. Все 10 ловчих моделей сосны, из которых 9 срублены в первой половине мая и одна в январе 1950 года, интенсивно заселились садовником и только две модели, выложенные в негоревших густых насаждениях, оказались слабо заселенными.
Стоящие усыхающие деревья успешно заселяются малым садовником только при достаточной степени ослабления и приостановки смолотечения. Эти условия создаются к весне в результате осенних пожаров или после заселения летом предыдущего года некоторых видов усачей.
Низовые пожары в спелых сосняках обычно не вызывают массового усыхания деревьев, но подчиненная часть древостоя слабее противостоит воздействию огня и постепенно в течение ряда лет погибает под натиском вредителей. Эти деревья и являются основной резервацией короедов на сосновых гарях.
Случается, что малый садовник нападает одновременно с усачами на здоровые стоящие деревья с хорошей кроной. Типичный пример — модель № 19 — 1950 года, диаметром 55 см, 220 лет, произраставшая в сосняке-брусничнике с полнотой 0,7 на южном склоне отрога вблизи «Четвертого столба». Садовник заселил эту сосну весной 1950 года совместно с усачами Callidium и Асаnthocinus carinulatus до высоты 7 м. К ним позднее присоединился в нижней части ствола Rhagium inquisitor. Средняя плотность по­селения садовника 9,2 маточных хода на 1 кв.дм, но большая часть этих поселений оказалась вместе с жуками без потомства, а маточные ходы залитыми смолой. На высоте 1 м ствола погибших поселений насчитывалось 47 проц. на высоте 3 м — 66 проц., на высоте 5 м, где оказалось наибольшее количество ходов усачей, всего 26 проц., а на высоте 7 м с полным отсутствием усачей — 98 проц.; в среднем на дереве погибло 62 проц. поселений. Эта модель, срубленная осенью 1950 года, имела густую зеленую крону, слегка начавшую желтеть в нижней части; там на сучьях были обнаружены поселения — Pityogenes irkutensis.
Отсюда вывод, что успешному размножению малого садовника на здоровом жизнеспособном дереве способствуют некоторые виды усачей, как первозасельщики. На стоящих деревьях именно усачи своими ходами сильнее нарушают физиологические функции дерева и этим способствуют последующему поселению других видов вредителей.
Биоценотические связи малого садовника на стоящих деревьях в районе горной тайги наблюдаются преимущественно со следующими видами. В нижней части ствола — Blastophagus piniperda, Hylurgops palliatus, Rhagium inquisitor, у шейки корня и на корнях Hylastes ater, Hylobius abietis; в средней части ствола В. minor, Acanthocinus aedilis, иногда Monochamus urussovi, а в предгорьях М. galloprovincialis. В этом комплексе обычен и Trypodendron lineatum. Верхняя часть ствола населяется обычно смолевкой — Pissodes pini.
Самые тонкие вершинные части стволов и ветви стоящих усыхающих сосен, а также ветви здоровых сосен, поврежденных климатическими факторами и усачами рода Monochamus, заселяются наиболее часто Pityogenes irkutensis, несколько реже — Р. bidentatus, еще реже Р. quadridens, изредка — Р. lichtensteini. Численность этих короедов возрастает с повышением в горы. Это — типичные виды горной тайги заповедника. На поваленных деревьях они не поселяются, по-видимому, вследствие быстрого пересыхания ветвей.
Биоценотические связи малого садовника на поваленных соснах примерно те же. В переходной полосе на высоте до 500 м к нему присоединяются преимущественно на поваленных деревьях Orthotomicus suturalis Gill и О. laricis Fabr. Встречаются они изредка и в горной тайге в сосняках-черничниках и вблизи долин ручьев. Здесь же найдены на повалах сосны и несвойственные ей виды — Hylurgops glabratus Zett и Drycoetes hectographus Reitt.
Отмирающий тонкомер сосны в сосняках-черничниках заселяется обычно в нижней части ствола В. piniperda, В. minor, Ortho­tomicus suturalis и О. laricis совместно с мелкими видами усачей — чаще из рода Leptura. В черничниках же и сосняках разнотравных переходной полосы встречается Carphoborus cholodkosky, но чаще этот вид заселяет сосновый тонкомер в сосняках-брусничниках. Blastophagus piniperda — большой садовник также относится к важнейшим видам вредителей, в частности на гарях, но значительно менее распространен и менее многочисленен, в силу меньшей экологической пластичности. Сроки его лета и всех фаз развития совпадают с таковыми малого садовника. Личинки его уничтожаются сильнее, чем у последнего, личинками серого длинноусого усача и хищников — Cleroides formicarius L, Pitho depressus и др., живущими также в нижней части стволов с толстой корой.
Фазы развития фиолетового лубоеда — Hylurgops palliatus Gill -, распространение которого отмечается преимущественно в тех же высотах, что и обоих садовников, укладываются почти в те же сроки, что и у двух предыдущих. Чаще гнездится в нижней части ствола, на пнях, выступающих корневых лапах. Отмечен случай довольно обильного заселения им совместно с Pissodes cembrae Mot свежезараженного кедра 80 лет из второго яруса на высоте до 9,5 м (модель № 70 на контрольной пробе № 2 в пихтарнике-зеленомошнике кисличнике). Местами фиолетовый лубоед встречается довольно часто, особенно на повале, но массовым вредителем не является.
Короедов — Pityogenes irkutensis и Р. bidentatus, особенно частых на сосновых гарях 1942 года Столбинского нагорья, следует отнести к массовым вредителям сосны. По нашим наблюдениям при некоторых обстоятельствах они в массе гнездятся на здоровых соснах, не способствуя, однако, их усыханию. Осенью 1949 и 1950 годов на свежесрубленных здоровых моделях сосны и лиственницы мы наблюдали массовое повреждение тонких ветвей диаметром до 1,0-1,2 см.
Живые ветви со свежей хвоей на расстоянии от конца 25-70 см на верхней стороне были как бы перерезаны бритвой на половину торца до сердцевины. Поверхность надрезанной части торца гладкая, словно отшлифованная, без признаков смолотечения. Надрез можно было заметить только при отгибе побега вниз. В.Н.Степанов, с которым автор консультировался по этому вопросу, связывает образование подобных поперечных трещин с резкими температурными колебаниями. Среди множества опавших под воздействием ветра сосновых и лиственничных ветвей, торцы которых были наполовину гладкими, наполовину с характерными особенностями излома, часть ветвей с признаками увядания были в массе заселены Р. irkutensis, P. bidentatus, иногда P. quadridens. Молодые жуки перезимовывают в опавших ветвях под снегом.
Одним из важнейших вредителей сосны — Ips sexdentatus Boern — стенограф встречен нами в большом количестве лишь в районе р.Слизневой на границе с Красноярским лесхозом, где этот вредитель на еловой гари 1945 года в нижней части склона совместно с большим и малым садовниками уничтожил полностью отдельные участки сосняков-брусничников. Стенограф отмечен и на других окраинах заповедника, примыкающих к районам лесозаготовок и сплава, в частности по р.Мане. На Столбинском нагорье, как исключение, найдены единичные поселения стенографа на северо-западном склоне к ручью Нелидовке близ просеки 31 и 48 кварталов в пихтарнике-зеленомошнике кисличнике, горевшем в 1942 году и основательно изреженном. Здесь найдены в 1950 году два свежих поселения по три маточных хода на комлях двух ветровальных деревьев ели и кедра.
Ips acuminatus Eichh — вершинный короед обнаружен в урочище «Ефтюгинские пашни» недалеко от р.Калтат. На Столбинском нагорье не обнаружен. По-видимому, стенограф и вершинный короед находят для себя наиболее подходящие условия в светлохвойных лесах предгорий, вблизи рек и ручьев, причем частота встречаемости их повышается по соседству с районами лесозаготовок. Центральная же горно-таежная часть заповедника с господством пихтовых лесов и островками сосняков не отвечает его экологическим особенностям.
Сроки лета Pissodes pini — сосновой столовой смолевки — очень растянуты. В 1949 году первый массовый лет был отмечен с 5 апреля по 20 июня и второй лет — с 11 июля по 30 июля. Налицо две фенологические популяции. Зимуют личинки.
На всех зараженных деревьях, где только поселяются малый садовник и смолевка, граница между поселениями тех и других всегда ясно выражена. Смолевка, откладывающая яички одновременно с садовником, или несколько позже, никогда не помещает свое потомство среди, хотя бы и очень редких, гнезд садовника. Селится выше поселений садовника, всегда в области тонкой коры. На кедре найден другой вид смолевки — Pissodes cembrae Mot со сходными биологическими особенностями развития. Смолевка является в заповеднике массовым вредителем, особенно на гарях, заселяя и стоящие и поваленные деревья. При дополнительном питании сильно вредит подросту сосны и кедра на гарях старше семилетней давности.
Лет большого соснового слоника — Hylobius abietis начинается в первых числах июня и продолжается в основном до два­дцатого июня. Слоник повреждает естественное возобновление сосны и кедра главным образом на старых гарях 15-25-летней давности и старше. На гарях 1942 года в сосняках-черничниках и брусничниках самосев сосны и кедра, возникший после пожара, слоником не повреждается.
По нашим наблюдениям долгоносик почти не трогает молодой слабый, этиолированный подрост сосны, произрастающий в группах пихтового и елового подроста, предпочитая подрост более крупный, достаточно хорошо освещенный.
Среди усачей наиболее опасным, в том числе и на гарях, является род Monochamus. M. urussovi — массовый вредитель всех хвойных пород, особенно пихты в горно-таежном поясе и пе­реходной полосе. M. galloprovincialis повреждает почти исключительно сосну. Район его деятельности — светлохвойные и смешанные леса предгорного пояса и частью переходной полосы.
Усачи рода Monochamus заселяют только совершенно свежие деревья, стоящие и поваленные, не заселенные другими вредителями или же слабо заселенные. В горной тайге лет жуков начинается со второй декады июля и особенно интенсивно в третьей декаде июля — первой декаде августа, заканчивается в первой половине сентября. Поэтому очень редко можно встретить личинок Monochamus на зимнем и весеннем ветровале и на срубленных в эти сроки деревьях, так как ко времени лета Monochamus они обычно уже заселены другими вредителями. Массовому размножению M. urussovi особенно способствуют весенние и летние пожары. Стоящие деревья он заселяет главным образом в нижней части ствола, но иногда и значительно выше. Особенно предпочитает пихту. Ветровальные и срубленные деревья пихты почти не заселяются в первый год другими вредителями, кроме M. urussovi, поскольку этому препятствует кора пихты, богатая смоляными ходами с долго не подсыхающей смолой.
В горно-таежном районе М. urussovi, имеет обычно двухлетнюю генерацию. В очень сырых и холодных условиях — на северном склоне в пихтарнике-зеленомошнике кисличнике нами наблюдались на поваленных затененных пихтах зазимовавшие двухлетние личинки М. urussovi, что объясняется неблагоприятными условиями погоды 1949 года. На достаточно свежем стоящем дереве нередко можно увидеть кроме двухлетних личинок также и однолетних.
М. impluviatus найдены единичные экземпляры на соснах в переходной полосе заповедника.
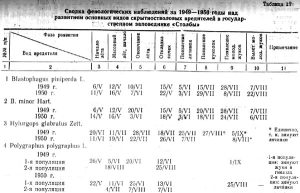
На сосне, а часто и на других хвойных породах, непременным обитателем нижней части ствола, а иногда и средней, является ребристый рагий — Rhagium inquisitor L, широко распространенный в обеих вертикальных зонах. Его личинки, а также личинки серого длинноусого усача своими широкими ходами часто совершенно истачивают внутренний слой коры, попутно уничтожая потомство большого садовника и фиолетового лубоеда. Сроки лета рагия очень растянуты (таблица 17). В 1949 и 1950 годах массовый лет его происходил с первой декады июня до двадцатых чисел июля, лет отдельных жуков — до осени. Зимовка отмечена на разных стадиях развития. Рагий размещает свое потомство особенно охотно на пнях, на нижней части стволов осенней рубки, на ветровале. Молодые личинки, как и личинки рогохвостов, постоянно встречаются на стволах деревьев, уже значительно подсохших, использованных другими вредителями в предыдущем году, однако довольно часто встречается и на живых деревьях всех хвойных пород, свежезараженных усачами рода Monochamus и другими вредителями.
Серый длинноусый усач Acanthocinus aedilis также предпочитает нижнюю часть ствола, но на поваленных деревьях заселяет и среднюю. Лет происходит ранней весной. Личинка его является одним из естественных врагов личинок обоих садовников, фиолетового лубоеда, иногда Orthotomicus laricis и О. suturalis. Acanthocinus carinulatus Gebl встречается преимущественно в переходной полосе и примыкающей к ней части горной тайги на стоящих, вполне жизнеспособных, но уже зараженных соснах, часто совместно с Callidium aeneum, иногда С. violaceum.
Asemum striatum L. встречается всюду часто, но не обильно, на всех хвойных породах.
Из других усачей очень широко распространены на сосне некоторые виды из подсемейства Cerambycinae родов Evodinus, Leptura и Strangalia, хотя имеют меньшее хозяйственное значение.
Из рогохвостов на сосне найден в массе только один вид — Paururus juvencus L, заселяющий в горно-таежном районе все хвойные породы. Другой, обычно конвергирующий с ним вид, — Paururus noctilio L встречен в небольшом числе в переходной полосе.
На пихте Abies sibirica Ledeb найдены только 5 видов короедов. Из них 4 вида сравнительно редко повреждают пихту. И только Xylechinus pilosus — пальцеходный лубоед, заселяющий также ель и кедр, является важнейшим массовым вредителем пихты, иногда кедра, на гарях всюду сопровождает темнохвойные породы, но заселяет только стоящие деревья.
На границе распространения пихты в долине ручья «Вторая Поперечная» при его впадении в ручей Лалетина, пальцеходный лубоед заселяет отдельные усыхающие тонкомерные ели, иногда заселяет совместно с Polygraphus subopacus усыхающие крупные ели. В районе горной тайги, в нетронутых пожаром насаждениях пихтарника-зеленомошника он заселяет часть отмирающего тонкомера ели и кедра, не заселенного гравером, а иногда и крупные кедры.
Зато на пихте это обычный вредитель, заселяющий совместно с усачами все естественно отмирающие деревья пихты. Так, на пробе № 2 в негоревшем пихтарнике-зеленомошнике этим лубоедом заселен на 91 проц. весь сухостой из главного полога, а имен­но 122 дерева на 1 га. Подрост пихты диаметром 4-8 см в ко­личестве 142 экз. на 1 га заселен им на 100 проц. Усачами эти деревья заселены на 85 проц., а подрост на 58 проц.
На гарях в пихтарниках пальцеходный лубоед размножается особенно бурно, начиная с 3-го года после устойчивого низового пожара. Возможно, он имеет две фенологические популяции, так как в одних случаях были найдены его поселения с зимующими личинками, в других случаях с зимующими молодыми жуками. В 1949 году в конце июля в его гнездах были найдены молодые жуки.
Лубоеды Hylurgops glabratus и Dryocoetes hectographus найдены были на пихте почти исключительно в негоревших насаждениях на поваленных деревьях.
Другой массовый вредитель пихты — Monochamus urussovi особенно опасен на гарях, так как, размножившись, держатся на гарях очень долго, постепенно заражая совместно с Xylechinus весь пострадавший в результате пожара древостой.
Семейство усачей, по сравнению с короедами, более широко представлено на пихте — ее заселяют не менее 24 видов усачей, причем большинство видов заселяет только стоящие деревья. Повалы пихты на гарях и вне их бедны видами. Их заселяют только усачи рода Monochamus, виды жуков Ernobius и др. Serropalpus barbatus L, древесинник Trypodendron lineatus. Видовой состав усачей на стоящих пихтах определяется особенностями фитоценозов, характером усыхания дерева, а также вредителями-доминантами данной биоценотической группировки.
Наиболее однообразный состав вредителей на стоящих деревьях наблюдается в загущенных негоревших пихтарниках-зеленомошниках кисличниках на северной экспозиции склонов: Monochamus urussovi, Rhagium inquisitor, Pogonochaerus, fasciculatus, Xylechinus pilosus, Trypodendron lineatus, Sirex gigas.
Усыхающий тонкомер плотно заселяется X. pilosus, некоторыми мелкими лептуринами, Pogonochaerus fasciculatus, Ernobius sp.
На гарях в этом типе леса видовой состав вредителей, вследствие изреживания древостоя, пополняется многими видами усачей из группы триб — Lepturites, Asemum striatum, Acanthocinus aedilis, Semanotus undatus и др., а также некоторыми рогохвостами, в том числе Xeris spectrum.
Одиночно стоящие, хорошо освещенные, усыхающие пихты исключительно богаты видами усачей, но короеды здесь отсутствуют. Кроме доминанта М. urussovi здесь отмечены М. saltuarius, Acanthocinus griseus, Pogonochaerus fasciculatus, Leiopus punctulatum, Asemum striatum, Clitus arietoides, Callidium violaceum, Rhagium inquisitor, Pachyta quadrimaculata, Evodinus borealis, Acmaeops septentrionis и др.; златки Ancilochira haemorroidalis, Ancilochira strigosa, Phaenops guttulata; Sirex gigas, Xeris spect­rum. не считая многих видов вредителей из не исследованных нами семейств.
Важнейшие виды короедов на ели Picea obovata Ledeb: типограф Jps typographus и матовый полиграф Polygraphus subopacus — весьма различны по своим экологическим свойствам.
Распространение первого связано с приручейными елово-пихтовыми насаждениями по долинам горных потоков в переходной полосе от нижнего горного пояса (предгорий) к горно-таежному. Вместе с елью по долинам рек проникает и в предгорья, а также языками и в горную тайгу, однако неглубоко и лишь отдельными поселениями. На еловых гарях, примыкающих к долинам рек, в ельниках-зеленомошниках в первые 2 года после пожара типограф размножается в массе, но после изреживания древостоя уступает свое место Polygraphus subopacus.
В 1949 году типограф приступил к устройству маточных ходов с конца мая, лет его был очень растянут. На некоторых деревьях кладки яиц наблюдались даже 18 июля. Окукливание, вследствие холодной дождливой погоды, началось только 20 июля, массовое окукливание — в первых числах августа.
Молодые жуки впервые замечены 1 августа, а в массе 15-18 августа. На многих деревьях молодое поколение осталось тут же на зимовку в ходах дополнительного питания, на отдельных деревьях с особенно высокой плотностью поселения и разрушенной корой, размокшей от частых осенних дождей, жуки вылетели в поисках более благоприятных мест для зимовки.
Обычная микростация типографа — весь ствол, за исключением вершины и ветвей, где поселяется гравер. В отдельных случаях на поваленных деревьях с очень высокой плотностью поселения он оттесняет гравера и занимает весь ствол полностью, а также толстые сучья (модель № 16 — 1949 г. в долине ручья Лалетина, недалеко от устья его притока — Второй Поперечной, модель № 8 −1950 г. у вершины увала горы «Пыхтун» в ельнике крупнотравном на склоне северной экспозиции). В заповеднике типограф охотнее заселяет поваленные, чем стоящие ели.
В противоположность типографу матовый полиграф заселяет исключительно стоящие деревья, причем хорошо освещенные. Но повал заселяет только высоко приподнятый над почвой. Обитает в пределах распространения ели и кедра, охотно заселяя и эту породу. Начало лета полиграфа отмечено в первой декаде июля. Единичные ранние жуки выводятся в третьей декаде августа, но в массе зимуют личинки.
Частый спутник матового полиграфа — пушистый полиграф селится обычно в нижней части ствола стоящих деревьев или на ветровальных елях. Имеет две фенологические популяции. Первую популяцию составляют жуки, зимовавшие в стадии imago. Лет их происходит в первой декаде июня, и они же часто выступают в роли первых засельщиков, вместе с усачами, ослабленных елей и кедров в нижней части ствола. Позже к ним присоединяется и матовый полиграф, заселяющий среднюю часть ствола. Вторая популяция зимует в стадии личинки. Лет ее на месяц позже первой, но растянут до середины августа. Особи этой популяции размещают свое потомство совместно с Р. subopacus, Hylurgops glabratus, Dryocoetes hectographus, причем с двумя последними гигрофильными видами заселяют преимущественно повал.
По ловчим модельным деревьям 1949 и 1950 годов видно, что крайней границей сплошного заселения ели типографом является в обследованном районе вершина горы «Пыхтун» вблизи кордона «Столбы» на высоте до 600 м над уровнем моря, то есть нижняя граница горной тайги. В пределах горной тайги отмечен спорадически в ограниченном количестве, причем на повалах ели уже не селится, полностью уступая место Н. glabratus с участием гектографа и пушистого полиграфа.
В противоположность пихте ловчие поваленные деревья ели заселяются усачами в очень ограниченном количестве. В биоценотических группировках с доминированием типографа такие основные виды, как Tetropium castaneum, Molorchus minor, Semanotus undatus, Callidium aeneum гнездятся главным образом на толстых сучьях совместно с долгоносиками — Magdalis frontalis, гравером и гектографом. В этой же группировке на толстых сучьях выведен нами и усачик Saperda interrupta Gebl, свойственный Восточной Сибири.
Более или менее обильное заселение ели усачами рода Моnоchamus наблюдается только на летнем и особенно ранне-осеннем ветровале, совпадающем повремени с летом Monochamus. M. sutor немногочисленен в заповеднике, но понемногу встречается всюду на темнохвойных породах. M. saltuarius Cebl встречается преимущественно в переходной зоне на ели и пихте, а в примыкающих предгорных борах на сосне и лиственнице.
На гарях в темнохвойных насаждениях на стоящих елях M. urussovi чаще встречается в сообществе с типографом, причем главным образом в первые годы после пожара, когда запас короедов еще недостаточен, а в древесине, годной для поселения, недостатка нет.
Наиболее типичными видами усачей на ели в заповеднике являются Tetropium castaneum и Semanotus undatus. В исследованном районе во всех высотных зонах на стоящих усыхающих елях наиболее частой биоценотической группировкой, во главе с обоими полиграфами, является следующая: нижняя часть ствола, особенно с лучше освещенной стороны, занята Tetropium castaneum, единично M. urrussovi; средняя часть ствола занята Callidium aeneum, Semanotus undatus, Molorchus minor и др.; вершинная часть и крупные сучья — Monochamus saltuarius, Pogonochaerus fasciculatus, Saperda interrupta, Magdalis frontalis.
Между поселениями усачей располагаются поселения короедов в пестром мозаичном порядке, но главным образом на лучше освещенном секторе ствола. Северный сектор ствола обычно сплошь занят усачами. Из короедов в этой группировке обычны оба полиграфа, гравер, изредка пальцеходный короед. К поселениям матового полиграфа часто примыкает свои ходы Crypturgus cinereus
В разных высотных зонах эта группировка вредителей варьирует по соотношению между отдельными видами.
На ветвях ели поселяются Pityophthorus micrographus, Pityogenes Saalasi, P. bidentatus, P. quadridens, Phthorophloeus spinulosus. Среди поселений гравера и матового полиграфа в районе Слизневой встречен Carphoborus rossicus Sem.
Тонкомер ели и кедра в горной тайге чаще всего заселяется гравером и Phthorphloeus spinulosus, в переходной полосе в районе ручья Лалетина, кроме них, также и X. pilosus, а в районе р.Слизневой Carphoborus rossicus Sem и Carphoborus teplouchovi Spess.
Ель — излюбленная порода для представителей сем. Siricidae. За исключением Paururus noctilio, найденного только на сосне, все остальные 5 видов, развивающиеся на хвойных, всеядны, но встречаются на ели чаще, чем на других породах.
На ели же найдены два уссурийских вида рогохвостов: Sirex umbra Sem и Paururus ermak Sem, из которых последний обнаружен в массовом количестве. Первый найден единично в переходной полосе, второй является часто массовым вредителем ели, иногда лиственницы в горной тайге и переходной полосе.
Из других семейств вредителей довольно часто встречается на поваленной ели технический вредитель Serropalpus barbatus (сем. Melandryidae).
Сибирской лиственнице Larix sibirica Lebed помимо обычных видов вредителей, свойственных другим хвойным, присущи свои специфичные виды: Jps subelongatus, Dryocoetes bajkalicus, Scolytus moravitzi, Tetropium gracilicorne, отмеченный нами единично также и на ели.
Наиболее существенное значение для лиственницы, как первые засельщики живых деревьев, имеют продолговатый короед Jps subelongatus и заболонник Моравица. Байкальский лесовик поселяется на деревьях уже заселенных другими вредителями и сильно ослабленных.
Тонкоусый дровосек Т. gracilicorne массовый вредитель лиственницы и заселяет ее одним из первых. Сначала он поселяется на нижней части ствола, иногда совместно с М. urussovi, на следующий год поселяется выше. Так как первоначальное его поселение происходит преимущественно на одной стороне ствола, более освещенной, то до окончательного усыхания дерева протекает иногда 3-4 года. Иногда дровосек занимает и свежеповаленные лиственницы совместно с М. urussovi и D. bajkalicus. Как и продолговатый короед, тонкоусый дровосек занимает доминирующее положение в образуемых ими группировках.
Заболонник Моравица заселяет верхнюю часть ствола, после чего к нему присоединяются другие вредители. Распространен главным образом в переходной полосе на высоте от 350 до 500 м, в районе ручья Вторая Поперечная.
Байкальский лесовик заселяет вершину, ветви и тонкомер лиственницы.
На лиственнице обычны также общие для всех хвойных серый длинноусый дровосек, ребристый рагий, Asemum striatum; Paururus juvencus, P. ermak, Phaenops guttudata Gebl.
На кедре Pinus sibirica Majr не найдено специфичных видов вредителей, за исключением смолевки Pissodes cembrae Mot. Кедр, произрастающий в насаждениях заповедника лишь в качестве небольшой примеси к другим, преимущественно темнохвойным породам, заселяется многими видами вредителей, свойственными и другим хвойным в зависимости от особенностей фитоценоза, в котором он произрастает.
Из короедов наиболее часто заселяется матовым и пушистым полиграфами, гравером, чернобурым и фиолетовым лубоедами, а в окраинных районах и стенографом. В негоревших пихтарниках-зеленомошниках кедр заселяется и пальцеходным лубоедом.
Из усачей кедру свойственны М. urussovi, Tetropium castaneum, Semanotus undatus, часто были находимы и личинки Callidium sр.
Серьезный вредитель кедра — смолевка Pissodes cembrae, заселяющая почти сплошь ослабленные, молодые сравнительно, кедры из второго яруса в пихтарниках.
По видовому составу вредителей, многообразию и сложности образуемых ими группировок кедр стоит ближе к ели, чем к сосне.
Из сказанного выше следует, что наиболее богата по видовому составу скрытностволовых вредителей переходная полоса от предгорий к горной тайге на высоте от 400 до 600 м, где перекрещиваются виды с самой разнообразной экологической требовательностью. Здесь проходит нижняя граница темнохвойных лесов, и эта же зона испещрена языками придолинных еловых, елово-пихтовых и смешанных лесов.
3. Динамика скрытностволовых вредителей леса на гарях
Гари заповедника как очаги заражения. Значению пожаров как очагов размножения скрытностволовых вредителей посвящено несколько специальных работ (С.С.Прозоров, 1929; В.И.Старк, 1925, и др.).
В работе С.С.Прозорова «Гари в сосновых лесах, как очаги заражения» отмечаются следующие наиболее существенные моменты в классификации гарей.
1. Время пожара: майские, июньские, августовские гари.
2. Характер пожара: беглый, низовой, устойчивый, низовой, верховой (или повальный).
3. Возраст насаждений, пройденных пожаром.
С.С.Прозоров считает наиболее опасным в отношении длительно действующего очага заражения устойчивый низовой пожар в приспевающих и спелых насаждениях.
Беглый пожар не отзывается существенно на древостое. Верховой пожар непосредственно приводит насаждение к отмиранию и почти исключает возможность заражения. Но устойчивый низовой пожар, обжигающий камбий у основания стволов, вызывает длительное болезненное состояние их, способствующее заселению вредителей и постепенному отмиранию деревьев.
Время пожара, по данным С.С.Прозорова, сказывается на видовом состоянии вредителей и последовательности заселения ими деревьев.
Результаты наших исследований на гарях заповедника, в основном Столбинского нагорья, полностью подтверждают эти закономерности с теми, однако, особенностями, которые присущи гарям в горных условиях, а также особенностями видового состава вредной энтомофауны и биоэкологии вредителей.
Одной из особенностей пожаров в горной местности является то, что на всех склонах и при всех условиях сильнее обгорают основание ствола и корни на стороне, обращенной вверх по склону. В этих поврежденных местах наиболее часто развивается гниль, особенно на пихте, и дерево вскоре переходит в категорию ветровала или бурелома.
Августовская гарь 1942 года занимает на Столбинском нагорье наиболее обширную площадь с разнообразным составом насаждений. Здесь были заложены 4 основные постоянные пробные площади в сосняках-черничниках и пихтарниках-зеленомошниках. В целом пожар можно охарактеризовать, как низовой, но в связи с разнообразием экологических условий (рельеф, экспозиция, состав древостоя, тип леса и проч.) сила огня и степень его воздействия на древостой и на почвенный покров также отличается разнообразием. Почти все насаждения гари представлены спелыми, частью приспевающими или перестойными сосняками, и в меньшей доле пихтарниками. Почти все эти насаждения подвергались и в прошлом неоднократным повторным пожарам. Совершенно не тронуты огнем на Столбинском нагорье лишь небольшие островки и сырые долины речек и ручьев.
Площади леса, подвергавшиеся в прошлом неоднократным пожарам, хотя и являются в настоящее время потухшими очагами массового размножения насекомых, все же отличаются от насаждений нетронутых пожаром несколько повышенным количеством мелких постоянных очажков заражения с высокой плотностью поселения вредителей. Степень близости этих постоянных очажков к площади свежей гари и видовой состав вредителей имеют большое значение для первоначального заселения каждой свежей гари.
Низовой пожар весны 1945 года в районе р.Слизневой захватил сосняки-брусничники верхней части юго-западного склона (обращенного к долине р.Слизневой), сосняки-черничники средней части, ельники-зеленомошники нижней части и шлейф склона. Наиболее пострадали от пожара насаждения верхней и нижней частей склонов. Первые — вследствие поверхностной корневой системы на мелких почвах с близкими выходами сиенитов, вторые — в силу неустойчивости к действию огня господствующей породы — ели. И те и другие насаждения явились очагами бурного размножения вредителей. Здесь были взяты две пробы в сосняках-брусничниках и две пробы в ельниках.
Беглый низовой пожар 1 мая 1948 года по восточному склону отрогов, спускающихся к р.Лалетина недалеко от северо-западной границы заповедника, охватил небольшую сравнительно площадь приспевающего насаждения сосняка разнотравного, в недалеком прошлом уже пройденного более сильным пожаром, повредившим камбий у многих деревьев на высоту до 8 метров. Несмотря на это, к 1949 году в насаждении усохла лишь небольшая часть деревьев из тонкомера, которая не может служить очагом размножения основных видов вредителей. Таблица 4.
Таблица № 4
Описание постоянных пробных площадей
Проба № 19 в кв. 48 около Музейного Камня. Гарь 1942 г. Тип леса Ptnetum myrtillosum. Древостой: 9С1Л+Ос - YI кл. возр. (120); ср. Д - 31 см. ср Н - 22 м; полнота 0,9: запас 321 куб. м. Второй ярус 10П+Е: ср. Д - 9,7; ср. Н - 10,0. Живой покров: восстанавливающийся покров черничника. Черника и другие боровые травы с обилием коротконожки. Пятна кипрея и венника. Нарушенный моховой покров с преобладанием Polyptichum sp.
Проба № 21 в кв. 31 около Караульного Камня. Гарь, 1942 г. Тип леса Pinetum myrtillosum. Древостой: 8С2Л+Ос - YII кл. возр.; ср. Д - 31: cp. Н - 19,5; полнота 0,9; запас 261 куб. м. 2-й ярус - 7П2Е1Б - полн. 0,1; ср. Д - 9; ср. Н - 7,5. Восстанавливающийся живой покров черничника. Черника, брусника и др. боровые травы. В нарушенном моховом покрове преобладает Polytrichum sp.
Проба № 24 в кв. 48. Южный склон за Вторым столбом. Гарь - 15-летней давности. Тип леса - Pinetum bracnvpodioso myrtillosum. Древостой: 9С1Л - VIII кл. возр. (150): ср. Д - 34: ср. Н - 24; полнота 0,8; запас 299 куб. м. В подросте редкие П.Е. В живом покрове коротконожка - Brachypodium pinnatum. Во 2 ярусе черника. Редкие пятна черничника-зеленомошника.
Проба № 16 в кв. 31. Урочище Барьеры. Гарь 1942 г. Тип леса - Pinetum vacciniosum. Древостой: 9С1Л - YII кл. возр. (140): ср. Д - 28; ср. Н - 19; полнота 1,0; запас 350 куб. м. Восстанавливающийся живой покров брусничника.
Проба № 3 (временная) в кв. 88. Верхняя часть юго-западного склона. Гарь 1945 г. Тип леса Pinetum vacciniosum. Древостой: 8С2Л - YII кл. возр. (135); ср. Д - 30; ср. Н - 23; полнота 0,85; запас 305 куб. м. Подрост - 10Е, редкий, ср. Д -11; ср. Н - 9. Живой покров густой из кипрея и Calamagrostis epigeios с примесью лесного разнотравья. Группы малины.
Проба № 1 в кв. 48, около избушки «Музеянка». Насаждение негоревшее. Тип леса - Рinetum myrtillosum. Древостой: 9С1Л+Ос+Б - YI кл. возр. (120); ср. Д -30; ср. Н - 23; полнота 1,0; запас 410 куб. м. Подрост - 4П4К2Е+С+Б средней густоты. Живой покров сплошной моховой с преобладанием мха Шребера. Небогатый травянистый покров с господством черники и примесью других боровых трав.
Проба № 20 в кв. 31. Сев.-западный склон к ручью Нелидовке, у просеки 31/48 кв. Гарь 1942 г. Тип леса Abiegnum hylocomioso-oxalidosum. Древостой: 7П2Б1С - YI кл. возр. (120): ср. Д - 16; ср. Н - 15,5; полнота 1,0; запас 261 куб. м; подрост средней густоты из П, Е, К. Живой покров из кипрея и вейников с сохранившимися пятнами типичной таежной растительности.
Проба № 22 в кв. 31. Западный склон хребта от камня «Щит». Гарь 1942 г. Тип леса Abiegnum hylocomioso-oxalidsum. Древостой: 7П2Б10с+Е+К - Y кл. возр. (100): ср. Д - 17,5; ср. Н - 16; полнота 0,7; запас 195 куб. м. Подрост средней густоты из П. Е. К. Живой покров из кипрея и вейников, густой, с редкими уцелевшими от огня пятнами типичной таежной растительности.
Проба № 2 в кв. 48. Северный склон к ручью Нелидовке, напротив избушки. Насаждение негоревшее. Тип леса Abiegnum hylocomioso-oxalidosum. Древостой: 7П1Е1К1Б - YI кл. возр. (120), ср. Д - 22: ср. Н - 19,5; полнота 0,9; запас 288 куб. м. Густой подрост из П. Е. К. Живой покров - сплошной моховой с преобладанием hylocomium-proliferum. Редкая кислица, теневые папоротнички и другие таежные виды.
4. Динамика энтомовредителей на гарях в сосняках
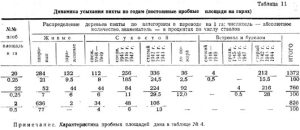
В сводной таблице 5 приведены данные по пяти наиболее типичным пробным площадям на гарях в сосняках и одной контрольной в негоревшем сосняке, а в таблице 4 дается описание этих проб.
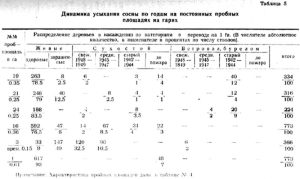
Из таблицы 5 видно, что наименее благополучные насаждения после пожара относятся к соснякам-брусничникам, особенно расположенным в верхней части склонов и на вершине увалов на мелких почвах с близкими выходами коренных пород. На пробе № 16 количество сухостоя достигает 23 проц. по числу стволов; на временной пробе № 3 — 91 проц. На гарях в черничнике — 16-20 проц. На контрольной пробе в черничнике — 10 проц.
В целом, за исключением пробы № 3, в сосняках-черничниках и брусничниках YI-YII классов возраста пожар не вызвал разрушения древостоя. Процент усохших и зараженных деревьев в течение 1949 года достигает на пробе № 19 — 4,5 проц.; на пробе № 21 — 13 проц. (только зараженные); на пробе № 24 (старая гарь) — 1,8 проц.; на контрольной пробе № 1 — 4,5 проц.; на пробе № 16 (брусничник) — 6,9 проц. Все эти деревья преимущественно из тонкомера. На пробе № 3 количество зараженных и усохших в 1949 году деревьев составляет 54,5 проц. Это насаждение на границе полного распада. Причина — мелкая почва с близким выходом сиенитов, повлекшая сильное повреждение огнем корневых систем. Имело значение и наличие в районе р.Слизневой такого опасного вредителя, как Ips sexdentatus Boern (см. таблицу 6). Повышенное количество зараженных деревьев на пробе № 21 объясняется более устойчивым характером низового пожара и сильным прогоранием оснований стволов, так как насаждение представляет собой вариант черничника, переходный к брусничнику, так назыв. бруснико-черничник.
На обожженной нижней части некоторых стволов после пожара поселились усачи из трибы лептурин, личинки которых устойчивы к смолотечению. Таких поврежденных живых стволов насчитывается 26 проц. На более устойчивых деревьях дальнейшего распространения усачей от зараженного места не произошло. Поэтому эти деревья в количестве 13 проц. мы отнесли условно к числу «здоровых». Остальные 13 проц., преимущественно тонкомерные, являются обреченными деревьями.
Обследование пробных площадей на Столбинском нагорье в 1950 году дало следующие результаты по сравнению с 1949 годом.
Из таблицы 7 видно, что в сосняках на гарях 1942 и 1934 годов ежегодно отмирает деревьев меньше, чем в негоревшем насаждении (проба № 1), где за отсутствием пожаров древостой гуще.

Одним из важнейших отрицательных последствий размножения на гарях лубоедов рода Blastophagus является, как известно, массовая подстрижка ими сосновых побегов.
На всех пробах, приведенных в таблице 8, процент деревьев с изреженными кронами очень высок, особенно в 1950 году. При анализе обстановки выявлено, что вблизи проб № 1, № 16, № 21 расположены (заложенные в 1949 году другим научным сотрудником) пробные площади по теме, предусматривавшей взятие большого количества модельных деревьев сосны. Модели, срубленные летом и осенью 1949 года, не были ошкурены и послужили в последующем резерватом для размножения обоих садовников. Участок насаждения, где заложена проба № 19, был неблагополучен и в 1949 году. По соседству с ним находится перестойный сосняк-брусничник с большим количеством усохших деревьев, зараженных садовником и Monochamus; довольно близко расположен участок пихтовой гари с обилием сухостоя и отмирающих деревьев, ставших рассадником усачей М. urussovi.

Для учета подстрижки крон осенью 1949 года на постоянных пробных площадях, заложенных в разное время на гарях и в негоревших насаждениях, были взяты 12 учетных площадок по 25 кв. м.
Наименьшее количество побегов, подстриженных большим и малым садовниками, оказалось на старой гари 23-летней давности в сосняке-черничнике (проба № 23), а именно 36 побегов в среднем на одной учетной площадке. На старой гари 1934 года (проба № 24) оказалось 74 подстриженных побега на учетной площадке. Особенно интенсивно происходила подстрижка крон на гари 1942 года. На отдельных площадках пробы № 19 и № 21 насчитано до 500 побегов, опавших в результате дополнительного питания молодых жуков садовника. Количество подстриженных побегов в негоревшем насаждении сосняка-черничника на западном склоне 4-го Столба (кв. 31) близко к таковому на пробе № 23.
Осень 1949 года отличалась сильными ветрами, и поэтому поврежденные побеги опадали в массе. Но в осенний сезон 1950 года сильных ветров не было. Часть поврежденных побегов осталась в кронах до более позднего времени и опадала уже по снегу, поэтому учет дал недостаточно полные результаты.
Так, на пробе № 19 на четырех площадках по мере приближения к соседнему негоревшему участку сосняка-черничника количество поврежденных сосновых побегов снижается: 52, 75, 19 и 11 побегов. В соседнем негоревшем участке сосняка-черничника, вдоль тропы, насчитано опавших поврежденных побегов: на расстоянии 30 м от гари — 12; 100 м — 0; 300 м — 39; 400 м — 52 и 430 м — 128. Последняя площадка была расположена вблизи неокоренных срубленных сосен. Дальше вдоль тропы от избушки «Музеянка» по направлению к р.Нелидовке на пробе № 1 на четырех площадках насчитано 52, 128, 92 и 62 побега. Поблизости от площадки с 128 побегами лежат остатки неокоренных моделей сосны, взятых при лесоустройстве в 1948 году. На площадке в центре пробы № 24 (сосняк-черничник) насчитано 19 побегов, на пробе № 21 на трех площадках от 1 до 4 побегов. Вверх по склону от этой пробы на трех площадках в сосняке-брусничнике насчитано 53,77 и на самой высокой точке 268 побегов. На пробе № 16 в сосняке-брусничнике в 1942 году найдено на одной площадке в центре пробы — 40 побегов. На свежеповаленных в октябре 1950 года соснах найдено, кроме того, большое количество побегов поврежденных садовником, но не опавших. Из этого следует, что способ учета повреждений садовником по опавшим побегам не отражает действительных размеров повреждений. Однако по соотношению повреждений на различных участках можно сделать заключение: 1) о прямой зависимости повреждений от сравнительной давности пожаров, причем, эти повреждения на гарях с течением времени уменьшаются; 2) о большей интенсивности повреждений на лучше освещенных и лучше прогреваемых деревьях и 3) о резком усилении повреждении на деревьях, ближайших к месту выплода молодых жуков в первичных очагах размножения.
На всех учетных площадках было найдено также большое количество побегов сосны и лиственницы размером до 0,5 м, опавших в результате воздействия климатических факторов, со следующей характерной особенностью: у всех побегов ровно на половине торца до сердцевины гладкая, блестящая поверхность без засмоления, а на другой половине торца следы излома, причем по расположению ветви на дереве излом приходится всегда на нижнюю сторону ветки, обращенную к почве. В 1949 году на отдельных учетных площадках пробы № 21 найдено было до 400 сосновых и до 50 лиственничных ветвей; среди них встречались единичные, пожелтевшие ветви, опавшие в результате повреждений молодыми жуками рода Monochamus с изломом на всей поверхности торца. В 1950 году из-за отсутствия сильных ветров, побеги, поврежденные в результате воздействия климатических факторов, составляли от 0 до 17 на площадку (преимущественно 3-4) и только в брусничниках на вершинах увалов, где ветер сильнее — от 37 до 41 на 25 м. Вместе с тем в кронах всех свежесрубленных осенью 1950 года соснах обнаружена масса не опавших побегов, поврежденных садовниками и климатическими факторами. Эти побеги густо заселяются короедами рода Pityogenes и Pityophthorus.
Соответствие между количеством побегов, опавших в результате повреждений лубоедами и в результате повреждений климатическими факторами, наблюдаемое на одних и тех же площадках, нуждается для своего объяснения в дополнительных исследованиях. Однако связь количества тех и других повреждений со сравнительной давностью гарей несомненна. Оба вида повреждений в наибольшем количестве отмечены на гарях 7-летней давности; несколько меньше их на гари 15-летней давности, значительно меньше на гари 23-летней давности и наименьшее количество отмечено в насаждениях не горевших или очень давно горевших. По-видимому, помимо прямого влияния пожара здесь имеет значение и косвенное — сравнительная осветленность сосняка, которая сильнее выражена в брусничниках и на молодых гарях с выгоревшим подростом и тонкомером. Кстати, сила ветра в таких светлых сосняках, особенно на вершинах увалов, больше, и это, несомненно, влияет на учет количества поврежденных побегов.
Экологические условия, создающиеся в области хорошо освещенных крон, с одной стороны, способствуют быстрейшему созреванию молодых жуков лубоедов, с другой стороны, благодаря резким сменам температуры и влажности ранней весной и поздней осенью, способствуют образованию глубоких поперечных трещин на молодых побегах.
На гарях в сосняках-черничниках через 15-20 лет при отсутствии повторных пожаров короедный запас иссякает, изреженные кроны оправляются, за исключением крон очень старых или больных деревьев. В сосняках-брусничниках гари содействуют сохранению на более длительное время высокой концентрации короедов, усиленному изреживанию крон (проба № 16), постепенному отмиранию ослабевших деревьев и изреживанию древостоя. Здесь наблюдаются случаи нападения короедов и на здоровые деревья (модель № 19, взятая на гари 23-летней давности). На вершинах увалов с особенно мелкой каменистой почвой устойчивый низовой пожар приводит иногда к почти полной гибели древостоя. Пример — проба № 3 в насаждении сосняка-брусничника на Слизневской гари 1945 года, где на 5-й год с момента пожара осталось только 9 процентов относительно здоровых живых деревьев, почти все с изреженными кронами. Одну из причин неустойчивости этого участка следует искать в структурных особенностях древостоя, прежде всего в высоком возрасте основной его части — 160-180 лет. Неустойчива и более молодая часть древостоя в возрасте 90 лет составленная из тонкомера, угнетаемого перестойными деревьями. Вторая причина, как уже упоминалось выше — произрастание на выходах сиенита с мелкими хрящеватыми почвами и, как следствие — сильнее повреждаемая пожарами поверхностная корневая система. И третья причина — нападение такого важного вредителя, как Ips sexdentatus.
По нашим наблюдениям, гари в сосняках-черничниках на Столбинском нагорье как очаги развития вредителей не оказывают заметного воздействия на прилегающие стены нетронутого огнем леса. Гораздо большее значение имеют первичные очаги в виде отдельных усыхающих зараженных деревьев и групп их. Пример — ряд площадок, взятых через некоторые промежутки вдоль Музеянской тропы в нетронутом огнем насаждении сосняка-черничника. Здесь, в непосредственной близости с гарью и на расстоянии до 300 м от нее, интенсивность стрижки невелика, затем резко возрастает в связи с первичным очагом размножения вредителей и далее к пробе № 1 опять постепенно затухает. В сосняках-брусничниках этого не удалось проследить из-за отсутствия по соседству с гарью участков брусничника, не тронутого огнем.
Интенсивность усыхания по годам, исчисляемая в процентах числа усохших в данном году ко всей живой части насаждения плюс сухостой данного года, приводится в таблице 9. В условиях заповедника интенсивность усыхания является одновременно и интенсивностью повреждений.

По данным таблицы 9 сильно повреждена проба 21 и особенно временная проба 3.
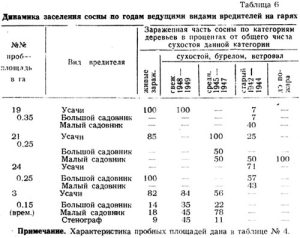
В следующей таблице 10 анализ зараженности вредителями сосны на гарях приводится по основным видам короедов; здесь же приводится объединенная группа усачей на сосне, в которой преобладают виды М. urussovi, Acanthocinus aedilis, Rhagium inquisitor.
Большинство обследованных деревьев заражено несколькими основными видами вредителей. Доминантом в образующихся здесь биоценозах является малый садовник, в меньшей степени большой садовник, фиолетовый лубоед, смолевка Pissodes pini, из усачей М. urussovi, Acanthocinus aedilis, Rhagium inquisitor, Callidium aeneum и Evodinus interrogationis.
Стенограф встречен, как главный вредитель, лишь на Cлизневских гарях.
Малый садовник дает почти одинаковую цифру заражения на всех пяти пробных площадях — около 42 проц. зараженных стволов сосны. Большой садовник более распространен на склонах к долинам рек и ручьев, в местах с повышенной влажностью воздуха (пробы № 24, № 3), где он заражает до 70 проц. стволов, в то время как в более сухих условиях (пробы № 19, № 21) заражает всего 9 проц.
Усачи дают на гарях самую высокую цифру заселения деревьев, особенно возрастающую на менее благополучных по степени повреждений гарях.
Динамика этих вредителей на гарях по годам представлена в таблице 6. Методика распределения прежнего сухостоя, бурелома и ветровала по годам повреждений дана выше. Особенности методики вытекают из того, что за неимением свежих гарей исследованием были охвачены гари в возрасте 4,7 и 15 лет.
На первых трех пробах, относительно слабо зараженных, с небольшим и неравномерным количеством сухостоя, трудно проследить закономерность заражения тем или иным видом по годам.
На Слизневской же гари эта закономерность хорошо проявляется. Многочисленная группа усачей дает на 4-й год после пожара наибольшую зараженность, которая, по-видимому, не снизится и в ближайшие годы. Все короеды дают наибольшую зараженность на 3-й год, после чего процент поврежденных ими деревьев от общего числа поврежденных в данном году сильно снижается. В отличие от В. piniperda и Jps sexdentatus, еще очень немногочисленных в первый годы после пожара, В. minor дает 78 проц. зараженных деревьев в первые же два года после пожара. Одновременно с нарастанием численности короедов резко возросла численность и их врагов, в частности наездников-браконид, а также других паразитных перепончатокрылых, и уже на 3-й год после пожара потомство стенографа при очень высокой степени зараженности было почти полностью уничтожено браконидами Jpobracon и Coeloides melanotus Wesm, после чего стенограф стал редким вредителем в данном районе.
Повреждения естественного возобновления сосны были обследованы нами на Столбинском нагорье.
На гарях 1942 года естественное возобновление еще совершенно не страдает от повреждений долгоносиками. Наблюдающаяся убыль самосева происходит, с одной стороны, вследствие этиоляции травянистым покровом в более влажных условиях местообитания и, с другой стороны, вследствие пересыхания почвы в более сухих условиях местообитания, особенно там, где пожар уничтожил моховой покров. На более старых гарях (пробы № 23 и № 24) долгоносики рода Hylobius и рода Pissodes являются опасными вредителями самосева сосны, кедра и других хвойных. Экологическая характеристика сравнительной повреждаемости самосева и подроста долгоносиками аналогична на обеих пробах. Основные выводы следующие:
1. В сосняках-черничниках наибольшая повреждаемость (на 100 проц.) подроста сосны и кедра, под кронами и у основания стволов деревьев. Скоплению в этих местах большого соснового слоника и смолевок способствовали, по-видимому, наиболее благоприятные для них условия защиты от прямого солнечного освещения и частых дождей, особенно в сезон 1949 года. Имело значение также и то, что к этим местам обычно приурочен подрост старших возрастных ступеней.
2. Большой процент поврежденного долгоносиками самосева от 60 до 83 проц. наблюдается и на небольших микровозвышениях, старых сглаженных выскорях.
3. На местоположениях по ровному склону с сильно прогоревшей подстилкой и покровом из послепожарных видов Polytrichum, лучше выраженном на пробе № 6, самосев повреждается относительно слабее, хотя количество поврежденного самосева доходит до 75 процентов.
4. В таких же микроусловиях, но со слабой степенью прогорания подстилки и быстро восстановившимся покровом из мхов Pleurozium Schreberi, Rhythidiadelphus triquetrus, Dicranum undulatum самосев повреждается долгоносиками значительно слабее и в количественном (около 30-40 проц.) и в качественном отношении.
5. Наименьшее количество поврежденного в слабой степени самосева (до 25 проц.) найдено на гребнях относительно свежих выскорей со слабо выраженным травянистым и моховым покровом, главным образом, из мелких лиственных мхов. Здесь же наблюдается и лучший рост самосева сосны.
Дополнительно нами был обследован самосев на небольшом участке сосняка-черничника, не тронутого пожаром, на юго-западном склоне к ручью «Столбинский Калтат» в квартале 31 вдоль просеки 31/48 кварталов. Насаждение состава 10 С+Л+Б+Ос YII класса возраста; средний диаметр 30 см, средняя высота 24 м, полнота 0,8; подрост пихты и ели высотой 3 м средней густоты. Моховой покров довольно мощный из мха Шребера с пятнами Dicranum undulatum, Hylocomium proliferum, Ptilium crista-castrensis. В травянистом покрове преобладает черника с примесью брусники и вейника тупоколоскового. Средняя высота подроста сосны — 32 см, средний возраст его 15 лет. Подроста, поврежденного слоником Hylobius abietis, оказалось 30 проц. Степень повреждения слабая.
Итак, наши наблюдения показывают, что в сосняках-черничниках на гарях 7-летней давности не наблюдается повреждений долгоносиками самосева и подроста хвойных пород; на гарях 15-летней давности эти повреждения носят массовый характер; на гарях 23-летней давности их несколько меньше; в насаждениях, не тронутых пожаром, или вернее, горевших очень давно, повреждений долгоносиками еще меньше.
Большой сосновый слоник повреждает периодически в массе самосев и подрост сосны и кедра; смолевки вредят естественному возобновлению всех хвойных.
5. Динамика энтомовредителей на гарях в пихтарниках-зеленомошниках кисличниках
В обследованном районе Столбинского нагорья пихтовые гари представлены только участками, горевшими в 1942 году. Динамика постепенного, в течение 7 лет, распада древостоя пихты показана в таблице 11. На пробной площади № 20 в квартале 31, где сила огня была относительно слабой, к осени 1949 года сохранился 21 проц. живых деревьев, к осени 1950 года их стало только 19,7 проц.
На пробной площади № 22, в том же квартале, соответственно сохранилось лишь 7,5 проц. и 6,6 проц. живых деревьев, причем очень ослабленных и интенсивно заселяющихся вредителями. Неустойчивость этого насаждения зависит от почвенных условий: близкие выходы грано-сиенитовых пород. Возникновению в этом месте пихтового древостоя способствовала узкая лощина, прорезавшая гриву с сосновым древостоем.
Наибольшее количество деревьев на пихтовых гарях погибло в первые 2 года после пожара: на пробе № 20 — 41 проц., на пробе № 22 — 68 проц. по числу стволов, не принимая во внимание деревья, усохшие до пожара. В дальнейшем усыхание происходило довольно равномерно: в среднем за год на пробе № 20 около 6 проц. древостоя, а на пробе № 22 около 4 проц. На контрольной пробе № 2 (квартал 48) в негоревшем насаждении в среднем за год отмирает около 2 проц. деревьев.
Основные массовые вредители пихты на гарях и вне их усачи рода Monochamus и пальцеходный короед — Xylechinus pilosus. Этим вредителям в негоревших пихтарниках свойственна очень высокая интенсивность заражения, высокая плотность поселения и энергия размножения.
Из таблицы 12 видно, что на гарях бывший сухостой до пожара сильно заражен Xylechinus. Резкое снижение процента зараженности им в первые два года после пожара объясняется тем, что наличного короедного запаса не хватает на большое количество деревьев, усыхающих сразу после пожара в результате непосредственного влияния огня.

Зараженность деревьев X. pilosus достигает кульминации в группе среднего сухостоя, то есть с 3-го по 5-й год, включительно после пожара, и затем снижается. Зараженность усачами на гарях непрерывно возрастала до времени исследования. На контрольной пробе не наблюдается закономерности повреждений.
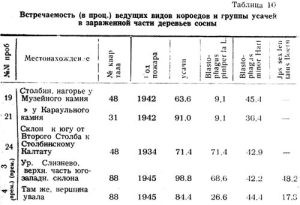
Влияние степени обгорания на зараженность можно проследить на отдельных группах деревьев каждой пробы. Приводим в качестве примера 7 площадок по 100 кв.м на пробе № 20 (таблица 13). Площадки выбраны по возможности с однородным травянистым покровом.
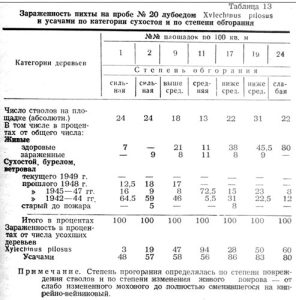
Заросли кипрея на площадках 1 и 2 указывают на сильное обгорание. В первые 2 года после пожара на этих площадках усохло около 60 проц. от всего сухостоя и бурелома. Здесь самая низкая зараженность усачами и X. pilosus.
На площадке 9 с покровом из редкого кипрея и вейника, где степень обгорания была несколько меньше, сохранился 21 проц. относительно здоровых живых деревьев. Из 46 проц. сухих деревьев почти половина использована лубоедом.
На площадке 11 со средней степенью обгорания в основной покров из сравнительно сохранившейся таежной растительности включены лишь пятна кипрея. Здесь усыхание деревьев происходило главным образом во втором периоде после пожара. Средняя группа сухостоя составляет 72,5 проц. и соответственно зараженность всего сухостоя лубоедом возрастает до 94 проц.
На площадке 17, несмотря на относительно слабое обгорание, при сохранившихся 38 проц. здоровых деревьев часть древостоя оказалась все же неустойчивой. Здесь преобладает группа старого сухостоя, процент деревьев зараженных лубоедом резко падает.
На площадках 19 и 24, с почти неизмененным пожаром живым покровом, зараженность лубоедом одна из самых высоких. На площадке 24, где в течение первых 5 лет после пожара усохло 20 проц. деревьев и дальнейшее усыхание прекратилось, зараженность X. pilosus составляет 60 проц.
Заражение усыхающих деревьев усачами находится в несколько меньшей зависимости от степени обгорания. При сильном и среднем обжигании ими заражено в среднем 55 проц. деревьев, при слабом — 83 проц. Это зависит от разнообразия видового состава объединенной нами группы усачей. Личинки одних мелких видов усачей, не определенных нами, были находимы только на небольших поврежденных огнем участках ствола совершенно здоровых в остальном деревьев. Но встречались также мелкие личинки усачей на деревьях усохших 6 лет тому назад. Основными видами усачей на пихтовых гарях являются Monochamus urussovi, Tetropium castaneum, Semanotus undatus, Rhagium inquisitor, некоторые мелкие лептурины.
На гарях динамика количеств поселений X. pilosus на пихте находится в зависимости от заселения последней М. urussovi.
По данным модельных деревьев старый сухостой довольно слабо заселен усачами и лубоедом, причем последний свободно селился два года подряд на одном и том же стволе до его окончательного усыхания. Наибольшим районом поселения лубоеда на стволе отличается сухостой среднего периода после пожара (до 50 проц. поверхности ствола). Но модели из сухостоя и зараженных деревьев 1948 и 1949 годов отличаются очень высокой плотностью поселения усачей и обширным районом их поселений — до 80 проц. поверхности ствола.
Поселения лубоеда здесь смяты ходами усачей и сохраняются отдельными небольшими участками.
Xylechinus pilosus не очень требователен к теплу и влаге. Модельные деревья показывают, что лубоед поселяется на всей окружности ствола, где только имеется свободная от усачей площадь. Однако чаще и плотнее заселяет стволы в более густых группах древостоя и менее охотно селится в изреженных участках леса.
Высота поселения для X. pilosus довольно безразлична. Часто заселяет ствол начиная с 0,4-0,5 м от шейки корня и до вершины, где его ограничивают другие вредители. Наибольшая плотность поселения лубоеда наблюдается на высоте ствола от 4 до 8 м.
По-видимому, уменьшение численности X. pilosus на гарях через 4-5 лет после пожара происходит как в силу повышения конкуренции усачей, так и вследствие изменения экологических условий после изреживания древостоя. Имеет также значение и возросший запас хищников и паразитов лубоеда.
6. Динамика энтомовредителей на гарях в ельниках-зеленомошниках
Нами обследована только одна еловая гарь 1945 года в районе р.Слизневой, в квартале 88. Пожар захватил на довольно большом протяжении правый крутой береговой склон к долине реки юго-западной экспозиции. В верхней части склона он захватил сосняк-брусничник, в средней части — сосняк-черничник, а в нижней части склона и по шлейфу — пихтарники-зеленомошники и ельники-зеленомошники, проходящие полосой в 100-150 м ширины по границе с заболоченной долиной реки.
Здесь в ельнике-зеленомошнике была заложена временная пробная площадь № 1 в 0,25 га, протянувшаяся на 100 м вдоль склона. Бывшая таксационная характеристика насаждения: 6Е2Л1Б+П+К; Y класса возраста; средний Д — 21,2 см, средняя Н — 21,0 м; полнота 0,7; III бонитет; запас 266 куб. м. В подросте редкие П, Е. Травянистый покров представлен главным об­разом кипреем Epilobium angustifolium, вейниками — Calamagrostis obtusata, C. Brachytricha, С. epigeios; местами заросли малины. Под ними сплошь Marschantia polymorpha, пятна послепожарных мхов Polytrichum sp, Funaria gigrometrica.
Анализ постепенного распада насаждения в результате пожара представлен в следующей таблице 14.
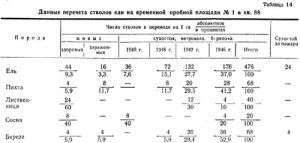
Распад елового древостоя происходит интенсивно. Через 4 года после разрушительного пожара осталось 9 проц. здоровой ели. Больше половины усохшей ели составляет ветровал. Пихта и береза пострадали еще сильнее. Усыхает постепенно и сосна.
Только лиственница, потерявшая в первые 2 года после пожара 40 проц. стволов, в дальнейшем не подвергалась усыханию.
Весь сухостой ели заражен вредителями. Динамика повреждений ее по годам представлена в таблице 15.

Кривая усыхания наглядно представляет структурные особенности распада насаждения по годам. В первую очередь под непосредственным воздействием огня усох весь тонкомер. Затем в ослабленном древостое под воздействием вредителей начинает отмирать господствующая часть. Наиболее крупный диаметр усыхающей части наблюдается на 3-й год (1948), после чего уменьшается. Средний диаметр оставшейся в 1949 году части древостоя снизился с 21,2 см до 16,5 см.
Зараженность усачами сухостоя на описываемой гари стопроцентная. Динамика зараженности короедами доминантами-типографом и матовым полиграфом представлена в таблице 16.
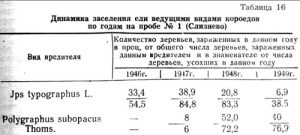
Типограф заселяет отмирающие и ослабленные деревья в первые два года после пожара, когда в насаждении еще имеются необходимые для него условия влажности и температуры. На третий год, когда деревья усохли (на 65 проц.) и более чем наполовину вывалились, на смену типографу являются виды более ксерофильные во главе с Р. subopacus.
С 1946 года типограф заселил 33,4 проц. общего количества заселенных им деревьев за весь период после пожара, причем с преобладанием деревьев слабо- и среднезаселенных. В 1947 году уже 38,9 проц., в том числе 20,8 проц. в сильной степени; в 1948 году только 20,8 ироц., преимущественно слабо заселенных; в 1949 году им заражено еще меньше — 6,9 проц. и тоже со слабой заселенностью.
Степень зараженности типографом определялась нами глазомерно с одновременным учетом района поселения и плотности поселения. За сильную степень зараженности условно принималось сплошное поселение типографа протяженностью не менее ½ высоты дерева с количеством маточных ходов от 5 и выше на 1 кв. дм поверхности коры. К средней степени относили зараженность ствола от ⅓ до ½ высоты и с плотностью поселения от 3 до 4 маточных ходов на 1 кв. дм. К слабо зараженным относили деревья с низкой плотностью поселения и расположением не выше ⅓ ствола.
Динамику заселения типографом по годам хорошо иллюстрируют цифры деревьев, зараженных в данном году, в процентах от суммы усохших за этот год. Так, в 1946 году типографом было заселено 54,5 проц. усохших в этом году деревьев. В 1947 году типограф заселил уже 84,8 проц., в 1948 году несколько меньше, а в 1949 году только 38,5 проц.
Размножение матового полиграфа происходит иначе. В первый год после пожара он отсутствует. На второй год, когда размножение типографа достигает наивысшей точки, матовый полиграф встречается всего на 6 проц. сухостоя этого года. На третий год он заселяет 72,2 проц. деревьев, а на четвертый год, когда типограф идет резко на убыль, 76,9 проц.
В процентах от всего количества деревьев, зараженных полиграфом, отдельно по годам имеем следующее. Появившись впервые в 1947 году, он заселил в слабой степени 8 проц. деревьев из числа хорошо освещенных. В 1948 году заселил 52 проц. деревьев, из них 32 проц. в сильной степени; в 1949 году — 40 проц., из них 28 проц. в сильной степени.
Глазомерный учет гравера — обычного спутника обоих предыдущих видов — провести не удалось, поскольку он селится в верхней части ствола. Обследование повала и модельных деревьев указывает на широкую распространенность гравера на этой гари за все годы после пожара.
Обычными спутниками типографа на поваленных елях является здесь также Dryocoetes hectographus (та повалах) и Hylurgops glabratus.
Обычными и широко распространенными спутниками матового полиграфа на стоящих деревьях являются Crypturgus cinereus и в меньшей степени Р. polygraphus.
Пихта на пробной площади заражена только усачами, главным образом М. urussovi на 77 проц. по числу усохших деревьев. Единичные деревья заселены в слабой степени X. pilosus. Этот лубоед, столь обычный на Столбинском нагорье, здесь в переходной полосе мало распространен.
Единичные кедры в насаждении усохли все в 1949 году, будучи заражены стенографом, гравером, Carphoborus rossicus, усачами не определенных нами по личинкам видов, а также Еrnоbius mollis — точильщиком.
Повал сосны в массе заражен М. urussovi, серым длинноусым усачем и рагием, Orthotomicus suturalis, реже О. laricis, Hylurgops palliatus, садовниками, а также Pissodes pini. Стоящие сосны поражаются усачами (чаще М. urussovi) и обоими садовниками.
Береза, усохшая почти полностью, повреждена на 56 проц. заболонником Scolytus ratzeburgi и усачами, отчасти древесинником Trypodendron signatus.
7. Значение климатических факторов в развитии скрытностволовых вредителей
Из фенологической сводки за 1949-1950 годы основных вредителей на Столбинском нагорье (таблица 17) видно, что отдельные фазы их развития не всегда совпадают по годам и протекают в зависимости от условий погоды данного года.
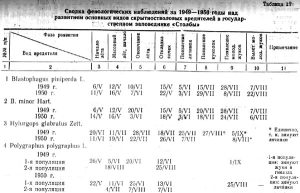
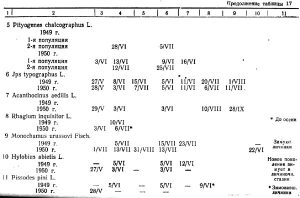
Лет ранних видов короедов — большого и малого садовников — в 1949 году начался 6 мая; массовый лет 12 мая, но холодная дождливая погода прервала его, и массовый лет происходил между 26 мая и 5 июня; закончился 10 июня. В 1950 году лет начался 11 мая; дружный массовый лет происходил в период между 16 мая и 29 мая. Неблагоприятная погода лета и начала осени 1949 года задержала развитие садовников на всех фазах. По сравнению с 1950 годом, погода которого может считаться близкой к средней за ряд лет по многолетним наблюдениям метеорологической станции на «Столбах», продолжительность массового лета в 1949 году затянулась на 12 дней. Появление первых куколок в 1949 году запоздало по сравнению с 1950 годом на 7 дней; появление молодых жуков — на 4 дня. Но и в 1950 году массовый вылет молодых жуков задержался на две недели, по-видимому, вследствие непрерывных дождей с 7 по 17 августа.
Состояние погоды отразилось, хотя и в меньшей степени, на короедах позднего лета, зимующих в стадии личинки. В 1950 году лет лубоеда Hylurgops glabratus начался на 9 дней раньше, чем в 1949 году; массовый его лет начался на 8 дней, а закончился на 3 дня раньше, чем в 1949 году. Появление молодых жуков гравера наблюдалось в 1950 году на 14 дней раньше, чем в 1949 году.
В 1950 году раньше начался лет и короедов среднего по времени лета, например типографа. Продолжительность же лета последнего в 1950 году составила 41 день, а в 1940 году — 50 дней.
В целом сроки лета и фазы развития насекомых в 1950 году наступили раньше и протекали дружнее, но в отдельных случаях развитие задерживалось периодическими дождями и наступившим в связи с этим похолоданием.
С другой стороны, по нашим наблюдениям, летние дожди способствовали вылету насекомых из-под коры, а длительная сухая погода задерживала вылет тех молодых насекомых, которые для высвобождения из древесины и коры пользуются размягчающим влиянием дождей. Летом после каждого дождливого периода наблюдалась обычно очередная волна массового появления разных групп скрытностволовых вредителей и их врагов, в частности наездников. В длительно-сухую погоду при осмотре коры хвойных мы часто наблюдали погибших вследствие застревания в коре и древесине усачей — рагия, серого длинноусого усача, лептурин и еще чаще наездников, особенно Xylonomus sp.
Нами наблюдалось влияние погоды на развитие вредителей, которая иногда даже вызывает их гибель.
В результате бурного развития осенью 1949 и 1950 годов подкорового грибка, обычного в сырых местах, погибло много личинок усачей в нижней части ствола различных хвойных пород. Грибок этот развивается в сырую погоду между корой и древесиной зараженных усыхающих деревьев, преимущественно с северной стороны. Мицелий грибка затягивает плотной белой пленкой лубяную часть коры от шейки корня на высоту до 1,5 м, включая корневые лапы; развивается также и на нижней стороне поваленных стволов. Определить его нам не представилось возможности.
Приводим следующие примеры гибели от подкорового грибка личинок.
1. На модели № 75 — стоящей ели в долине р.Лалетина, на высоте ствола до 1 м, погибло 85 проц. личинок Теtropium castaneum, Rhagium inquisitor, Semanotus undatus и др.
2. На модели № 22 — 1950 года стоящей лиственницы возле избушки «Музеянка» в сосняке-черничнике, зараженной Tetropium gracilicorne, но еще живой, на северном секторе ствола до высоты 1,5 м, личинки усача погибли на 90 проц., а личинки браконида Atanycolus initiator Nees в коконах погибли на 22 проц.
Вследствие холодной погоды 1949 года около 40 проц. а в отдельных случаях до 60 проц. личинок усачей Monochamus, Tetropium, Semanotus не достигли достаточного развития и не смогли углубиться в древесину до наступления заморозков. Это сделало их легко доступной добычей для дятлов, а также вызвало заболевание личинок.
Из обследованных зимой 1949-1950 годов под корой сосен личинок М. urussovi, взятых в различных экологических условиях, 18 личинок (или 37 проц.) оказались потемневшими, вялыми. Весной 1950 года на некоторых моделях сосны, ели и пихты среди находившихся под корой перезимовавших личинок Моnоchamus, и в меньшей степени личинок Rhagium Tetropium и др., оказалось от 30 до 35 проц. личинок, погибших от неизвестной причины.
Как мы указывали выше, погода оказывала влияние и на глубину расположения куколочных колыбелек у обоих садовников.
Климатические особенности года сказались на видовом составе и встречаемости отдельных групп насекомых-хищников и паразитов, проживающих под корой.
Группа жуков-хищников, как например, Cleroides formicarius, Rhizophagus dispar, Glichrochilus quadrimaculatus, Hypophloeus suturalis и др., весьма многочисленная в 1949 году, в следующем году резко снизилась в своей численности. Так, на моделях сосны, взятых в 1949 году, личинки Cleroides formicarius в ходах садовников составляли 0,5 экз. в среднем на 1 кв. дм; в 1950 году на многих моделях они совсем отсутствовали, а на остальных личинки Cleroides составили в среднем лишь 0,1 экз. на 1 кв. дм. Зато мелкие стафилиды и их личинки, энергично уничтожавшие яички и мелких личинок короедов, наблюдались в 1950 году в значительно большем количестве.
Наездники хальциды прекрасно развивались в 1949 году на стоящих деревьях, на поваленных же были немногочисленны.
Чрезвычайно многочисленны были в 1949 году как на сосне, так и на ели личинки двукрылых. В 1950 году они наблюдались во множестве на повалах ели в ходах типографа, но на сосне и стоящей и поваленной их было мало, зато на поваленных соснах в ходах малого садовника личинок хальцид и браконид было больше по сравнению с 1949 годом.
Горный ландшафт заповедника с типичными для него относительно небольшими высотами (в основном до 700 м над уровнем моря) отличается изрезанностью рельефа и поэтому исключительным разнообразием экологических условий на относительно небольшом пространстве. Отсюда одной из основных особенностей влияния горного климата заповедника на энтомофауну является чрезвычайная растянутость лета даже таких вредителей, как большой и малый садовник.
Эти особенности фенологии развития вредителей затрудняют установление единых твердых сроков для мероприятий по борьбе с вредителями. Сроки надо установить применительно к экологическим условиям и разным популяциям.
8. Мероприятия по борьбе с вредителями
Основные виды скрытностволовых вредителей лесов заповедника, с которыми надлежит проводить меры борьбы, следующие: Blastophagus minor Hart, Jps typographus L., Polygraphus subopacus Thoms, Xylechinus pilosus Ratz, Pityogenes chalcographus L, а по окраинам заповедника также Jps sexdentatus Boern; из усачей Monochamus urussovi Fisch, M. galloprovincialis var. pistor Cerm, Tetropium castaneum L, Tetropium gracilicorne Reitt. Остальные виды, входящие в биоценотические группировки, образуемые перечисленными выше доминантами, будут уничтожаться попутно.
Борьбу с вредителями в условиях хозяйства заповедника возможно проводить в следующих направлениях:
1. Лесохозяйственные профилактические мероприятия.
2. Лесохозяйственные мероприятия по борьбе с вредителями в постоянных очагах размножения.
3. Мероприятия по борьбе с вредителями в очагах массового размножения вредителей.
Особенности заповедного режима не позволяют рекомендовать многие настоятельно необходимые мероприятия по борьбе с вредителями. Но и в тех случаях, когда сильные пожары приводят к гибели всего насаждения, к массовому размножению вредителей и настоятельно необходима своевременная вырубка обреченного древостоя, — это часто не является возможным из-за отсутствия путей транспорта и сбыта заготовленной древесины.
Наиболее опасными и длительно действующими очагами массового размножения являются в заповеднике гари в темнохвойных насаждениях и в сосняках-брусничниках, расположенных на мелких почвах с близкими выходами коренных пород, особенно в спелых и перестойных насаждениях.
К профилактическим относятся противопожарные мероприятия, а также меры, обеспечивающие нормальное состояние леса с восстановлением его коренных типов, улучшение состава, в частности, восстановление таких ценных долговечных пород, как кедр и лиственница, и уменьшение роли пихты в лесах заповедника. К этой цели ведут такие лесохозяйственные мероприятия, как рубки ухода, осветление подроста ценных пород, уход за естественным возобновлением на гарях и его видовым составом, реконструкция малоценных молодняков на старых вырубках, содействие естественному возобновлению в изреженных пожарами перестойных насаждениях, а в отдельных случаях и культуры ценных пород.
Мероприятия второй группы имеют назначением ликвидацию первичных так называемых постоянных очагов размножения. В исследованном нами районе наибольшая концентрация первичных очагов наблюдается на гарях. Здесь должны применяться санитарные рубки, а также своевременная разработка и окорка ветровальных и буреломных деревьев.
Санитарные рубки следует в заповеднике планировать в первую очередь на площадях всех гарей давностью до 10-15 лет. Сюда же можно включить и свежие гари, возникающие в сосняках всех типов, пройденных низовым пожаром, за исключением некоторых брусничников на мелких каменистых почвах, состояние которых безнадежно и, возможно, потребуется сплошная их рубка.
При невозможности для заповедника охватить санрубкой всю территорию заповедника следует запланировать под санрубку наиболее неблагополучные участки вблизи особо ценных насаждений или же отдельные районы заповедника, в частности экскурсионно-туристический, ограничиваясь вырубкой только свежезараженных деревьев.
Отбор и клеймение деревьев, зараженных вредителями раннего лета (оба садовника, типограф, гравер ранней фенологической популяции и др.) следует производить во второй половине мая, когда произойдет заселение; рубку же и окорку — во второй половине июня, до окукливания и вылета жуков. Отбор и клеймение деревьев, зараженных вредителями среднего и позднего лета, каковы Р. subopacus, X. pilosus, усачи рода Мonochamus и Tetropium, зимующие в личинках, производить в первой половине августа (для Monochamus несколько позже), а вырубку и окорку во второй половине августа, не позже первой декады сентября.
При отсутствии возможностей к использованию древесины на площади ветровала, а также при единичном и групповом ветровале необходимо произвести окорку такового в порядке лесохозяйственных работ в надлежащие сроки.
Рекомендуем следующие сроки окорки стволов, зараженных основными видами вредителей в условиях заповедника «Столбы»:
Для В. minor u В. piniperda с 20/YI по 10/YII
« Jps typographus с 15/YI по 15/YII
« Polygraphus subopacus с 15/YII по 10/YIII
« Pityogenes chalcographus 1-я популяция с 15/YII по 30/YII
2-я популяция с 30/YII по 15/Y следующего года
« Xylechinus pilosus с 30/YIII по 15/Y
« Jps sexdentatus с 20/YI по 15/YII
« Monochamus urussovi с 1/YIII по 1/IX
« Tetropium castaneum с 15/YI по 15/YIII
По возникновении свежих гарей вопрос о назначении хозяйственных мероприятий разрешается на основе учета всей совокупности условий пожара и экологической обстановки.
В сосняках-черничниках низовые пожары отзываются почти исключительно на тонкомере из отставших в росте деревьев и из деревьев более молодых поколений девственного разновозрастного леса. Тонкомер сильнее повреждается огнем и в течение нескольких лет постепенно усыхает под влиянием вредителей. Здесь планируются только санитарные рубки с соблюдением сроков окорки.
В сосняках-брусничниках на почвах очень мелких каменистых, как уже упоминалось выше, после устойчивого низового пожара возникает необходимость в вырубке всего древостоя. Безнадежными деревьями следует здесь считать такие, у которых корневая шейка и корневые лапы повреждены более чем на 50 проц. При наличии таких деревьев свыше 60 проц. в полном насаждении — насаждение приходится назначать в сплошную рубку. При относительно благополучном состоянии древостоя планируется санитарная рубка. Беглый пожар в сосняках почти не имеет значения для размножения вредителей.
В темнохвойных насаждениях, особенно пихтовых, низовые пожары создают длительные устойчивые очаги размножения вредителей с постепенным отмиранием всего древостоя. В таких насаждениях при возможности вывозки и использования древесины следует назначать сплошную рубку. Разработка должна быть проведена в первый же год после пожара с обязательной окоркой древесины. При невозможности использования древесины проводятся санитарные мероприятия по своевременной окорке зараженных стволов.
Майские пожары в сосняках заповедника не влекут за собой немедленного заселения короедами раннего лета, каковыми являются садовники и стенограф. Они поселяются обычно на второй год после пожара. Но запоздавшие особи частично могут поселиться и в том же году. В год пожара на ослабленных деревьях поселяется в августе Monochamus. Июльские и августовские гари могут заселяться в тот же год только Monochamus.
Майские гари в пихтарниках могут заселиться в тот же год пальцеходным лубоедом, особенно при одновременном заселении усачей.
Биологический метод борьбы с вредителями должен найти особенно широкое применение на гарях в горных лесах заповедника, трудно доступных для разработки и вывозки древесины. Его следует проводить в двух направлениях: 1) привлечение полезных птиц, уничтожающих личинок и жуков, 2) содействие усиленной концентрации на гарях насекомых, являющихся врагами вредителей.
Среди птиц наибольшее значение в уничтожении скрытностволовых вредителей на гарях заповедника имеют: восточная желна, сибирский большой пестрый дятел, обыкновенный поползень, сибирская обыкновенная пищуха. Для привлечения дятлов обычно сохраняются на гарях и по окраинам их осиновые деревья. Дуплянки для них не имеют значения.
Для увеличения численности полезных насекомых, являющихся врагами вредителей леса, следует оберегать их потомство. Поскольку цикл развития наездников часто не совпадает с таковым их хозяев, при проведении санитарных мероприятий следует строго учитывать сроки развития тех и других. При запоздании в сроках окорки, если основные вредители уже вылетели, окорку производить не следует, так как под корой остаются личинки и коконы наездников и двукрылых, imago которых появится только в июне следующего года. Следует учесть, что наибольшее количество наездников развивается на стоящих, хорошо освещенных деревьях.
Запас наездников создается в лесу всюду, где имеется запас их хозяев-вредителей. Чем неразборчивее паразит при выборе хозяина, тем выше его численность и шире в тоже время экологические условия, в которых он встречается.
При достаточном запасе паразитов и хищных насекомых в соседних с гарью насаждениях, особенно имея в виду, что большинство наездников по сравнению с многими короедами плохие летуны, — весьма желательно выпускать на гари дополнительное количество наездников-браконид, а также хальцид. Для этого весною на стоящих деревьях сосны и ели, использованных садовниками и типографом в предыдущем году, можно собрать куски коры с кокончиками паразитов и перенести на гарь. Кору развесить в защищенных местах, сложив ее пачками по 2-4 куска, приложив внутренней стороной с коконами друг к другу. Надо стараться сохранить их от птиц и муравьев. Коконы наездников можно начать собирать еще с осени и хранить до весны в прохладном помещении.
Literature
1. Арнольд и Л.В. Жесткокрылые или жуки. «Животный мир СССР», т.4, изд. АН СССР, М.-Л., 1953.
2. Белановский И.Д. Закономерности в массовых размножениях вредителей в связи с метеорологическими факторами. Зоологич. журн., т.XY, в.2. 1936.
3. Белоусов В. Короеды Северных Саян. Энтом. обозр. XYI. № 3-4, СПБ. 1916.
4. Буковский В.О. О некоторых принципах организации энтомологических исследований в заповедниках. Н.-методические записки Гл. Управл. по заповедникам, вып. XI, М., 1948.
5. Гусев В.И. и Римский-Корсаков М.Н. Определитель повреждений лесных и декоративных деревьев и кустарников Европ. Части СССР. Изд. 3-е, М.-Л., 1951.
6. Гуссаковский В.В. Рогохвосты и пилильщики. Фауна СССР, сер. 1, т. II, вып. 1, М.-Л., 1935.
7. Зиновьев Г.А. О структуре, динамике и типологии очагов размножения короедов. Зоологич. журнал, т. XXXYII, вып. 3, 1958.
8. Ильинский А. Закономерности в размножении малого соснового лубоеда и теоретические обоснования мер борьбы с ним в лесу. «Защита растений», т. Y, М., 1958.
9. Ильинский А. Определитель яйцекладок, личинок, куколок насекомых. М.-Л., 1948.
10. Киселева Е.Ф. Короеды Томской области. Тр. Томского Гос. университета, 97, Томск, 1946.
11. Коротнев Н.И. Короеды, их лесоводственное значение и меры борьбы. Экология короедов Восточной Европы, Кавказа и Сибири, М., 1926.
12. Куренцов А.И. Экологические группировки ипидофауны (короедов) в связи с типами леса. Вести. Дальневост. фил. АН СССР, 9 М., 1934.
13. Куренцоа А.И. Короеды Сихотэ-Алинского заповедника. Тр. Сихотэ-Алинского гос. заповедника, II, М., 1938.
14. Куренцов А.И. Короеды Дальнего Востока СССР. Изд. АН СССР. М.-Л., 1941.
15. Лавров С.Д. К энтомофауне предгорий Саян. Тр. Сибирск. сельскохоз. акад., IY, 7, 1926.
16. Медведев С.И. Жесткокрылые. «Животный мир СССР». III. Изд. АН СССР. М.-Л., 1950.
17. Определитель насекомых Европейской части СССР, ОГИЗ, М., 1940.
18. Осипов А.И. Борьба с короедами в насаждениях сосны и ели. М., 1938.
19. Плавильщиков Н.Н. Жуки-дровосеки вредители древесины. Гослестехиздат, М.-Л., 1932.
20. Плавильщиков Н.Н. Жуки-дровосеки, ч.1. Насекомые жесткокрылые, т. XXI, Фауна СССР (нов. серия), изд. АН СССР, Л., 1936.
21. Плавильщиков Н.Н. Жуки-дровосеки, ч. 2. Насекомые жестко­крылые. т. XXII. Фауна СССР (нов. серия). Изд. АН СССР, Л., 1940.
22. Плавильщиков Н.Н. Жуки-дровосеки, ч.3. Фауна СССР, т. XXIII. вып. 1. Изд. АН СССР, Л., 1958.
23. Положенцев П.А. Биологические эксперименты и наблюдения над усачом Monochamus galloprovincialis Oliv на ветровале. Н.-методические записки Гл. Управл. по заповедникам, вып. XI, М., 1948.
24. Правдин Ф.Н. Программа работ по теме: изучение закономерностей массового размножения насекомых — вредителей заповедных лесов. Н.-методические записки Гл. Упр. по заповедникам, в. XI, М., 1948.
25. Прозоров С.С. Гари в сосновых лесах, как очаги заражения. Труды Сибирского ин-та с/хоз-ва и лесоводства, т. XII, вып. 1-3. Омск, 1929.
26. Рихтер А.А. Златки. Фауна СССР, XIII, 2. М., 1949.
27. Семенов-Тян-Шанский А.П. Пределы и зоогеографические подразделения палеарктической области для наземных сухопутных животных на основании географич. распределения жесткокрылых насекомых. Изд. АН СССР, М., 1936.
28. Семенов-Тян-Шанский А.П. Жуки. «Животный мир СССР», т.1, М., 1936.
29. Сокановский Б. Заметки по систематике и географическому распространению палеарктических короедов. «Защита растений», Y. М., 1928.
30. Сокановский Б. О биологической пластичности короедов и факторах ее развивающих. «Защита растений», Y, М., 1928.
31. Спесивцев П. Определитель короедов Европейской части СССР (за исключ. Крыма и Кавказа). СХГИЗ, М.-Л., 1936.
32. Старк В.Н. К типологии короедных очагов хвойных Карачижско-Крыловской лесной дачи Брянской губ. «Защита растений», II, М., 1925-а.
33. Старк В.Н. Значение пожарищ в деле образования короедных очагов в Брянской губ. «Защита растений», II. М., 1925-б.
34. Старк В.Н. Образование короедных группировок в зависимости от полноты насаждений. «Защита растений», т. Y, в. 3-4, М., 1929.
35. Старк Н.К. Об энтомофауне типов леса «Защита растений», YI, М., 1930.
36. Старк Н.К. Враги леса. Изд. 2-е, Сельколхозгиз, М.-Л., 1931.
37. Старк В.Н Вредные лесные насекомые. Сельколхозгиз, М.-Л., 1931.
38. Старк В.Н. Руководство к учету повреждений леса. Сельхозгиз., изд. 2. М.-Л., 1932.
39. Старк В.Н. Короеды. Фауна СССР, новая серия № 49, т. XXXI, Изд. АН СССР, М.-Л., 1952.
40. Степанов В.Н. К вопросу о насекомых — вредителях лиственницы сибирской на Южном Урале. Труды Ильменского госзаповедника им.В.И.Ленина, вып. IY, ОГИЗ. Челябинск, 1949.
41. Тальман П.Н., Яцентковский А.В. Вредные насекомые еловых и елово-лиственных лесов и меры борьбы с ними. Гослестехиздат, М.-Л., 1938.
42. Тальман П.Н. и Носырев В.Н. Метод массового детального обследования древостоев, зараженных вторичными вредителями. Н.-методич. записки Гл. Управл. по заповедникам, вып. XI. М., 1948.
43. Флеров Б.В. Повреждаемость возобновления сосны большим сосновым слоником и необходимость борьбы. Записки лесной опытн. станции при Тихвинском леспромхозе, IY. Л., 1931.
44. Флоров Д.Н. Насекомые — вредители хвойных насаждений Восточной Сибири. Иркутское областное изд-во, 1938.
45. Флоров Д.Н. Короеды хвойных деревьев Восточной Сибири. Иркутское областное изд-во, 1949.
46. Флоров Д.Н. Вредители сибирского кедра. Иркутское обл. изд-во, 1951.
47. Флоров Д.Н. Жуки-смолевки хвойных деревьев Вост. Сибири. Изд. БТНИИ при Вост.-Сибирском ун-те, т. X, вып. 4. Иркутск.
48. Шипулин А.Я. О сроках окорки древесины пихты сибирской. Лесное хоз., № 4, 1950.
49. Яцентковский А.В. Определитель короедов по повреждениям. Сельхозгиз, М.-Л., 1930.
50. Яцентковский А.В. Главнейшие вредные насекомые в лесах СССР. Сельколхозгиз, М.-Л., 1931-а.
51. Яцентковский А.В. Обследование короедников. Сборник статей Ленинград. лесотехн. академии. «Обследование лесов, зараженных вредителями». Л., 1931-б.
Owner →
Offered →
Collection →
Севастьянова Татьяна Петровна
Севастьянова Татьяна Петровна
Труды государственного заповедника