Труды государственногозаповедника "Столбы". Выпуск III. Насекомые паразиты и хищники скрытностволовых вредителей на гарях государственного заповедника «Столбы»
Введение
Наряду с изучением скрытностволовых вредителей леса заповедника, результаты которого изложены в предыдущей статье «Скрытностволовые вредители леса на гарях государственного заповедника «Столбы», в 1949-1950 годах изучались и их естественные враги — насекомые паразиты и хищники из отрядов Coleoptera, Hymenoptera, Hemiptera и др. Эти материалы и составляют содержание настоящей статьи.
Изучение деятельности других естественных врагов скрытностволовых вредителей, а именно птиц, не входило в нашу задачу. По этому вопросу автор располагает лишь следующими небольшими наблюдениями.
Наиболее часто и в больших количествах истребляются птицами, в частности дятлами, личинки короедов, смолевок и тех видов усачей, личинки которых или зимуют под корой, или сравнительно неглубоко проникают в древесину, например, личинки Tetropium grас licorne. Здесь дятлы извлекают также закоконировавшихся личинок, особенно наездников Atanvcorus maculator F. Xorides colearis Grav и др.
Из короедов особенно часто подвергаются истреблению дятлами, поползнем и др. личинки лубоедов Н. giabritus D. hestograpius P. poiy raphus, зимующие в стадии личинки. Осенью, когда личинки достигают больших размеров и так сильно истачивают кору, что она легко снимается, птицы часто полностью вскрывают кору и истребляют все потомство этих лубоедов. В меньшей степени истребляются короеды раннего лета, успевающие к осени пройти весь цикл своего развития.
Изучение насекомых — хищников и паразитов скрытностволовых вредителей — автор занимался главным образом в 1950 году, после оборудования на Столбинском нагорье энтомологической лаборатории с разнообразными садками для выведения насекомых.
Общее количество собранных и выведенных автором видов паразитических перепончатокрылых составляет 53 вида, в том числе из подсем. Jchneumonidae 25 видов, подсем. Braconidae 19 видов, сколий 1 вид и мелких видов перепончатокрылых из подсем. Chalcididae и других 8 видов. Кроме того, собрано 15 видов жуков-хищников, 15 видов хищных двукрылых, несколько видов хищников из других отрядов насекомых.
Из этих сборов нами определено (см. таблицу 1) 12 видов жуков-хищников, 39 видов паразитических перепончатокрылых, 8 видов хищных двукрылых, 1 хищный клопик, 1 вид сколии, 2 вида жуков-сожителей короедов.


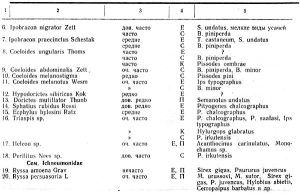
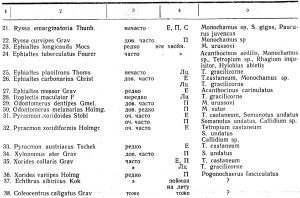

Из числа собранных наездников паразитами короедов являются 8 видов браконид, 2 вида ихневмонов, кроме того, 8 видов хальцид, 1 вид сколий (Tiphia minuta Lind), а всего 36 проц. найденных видов. На усачах, рогохвостах и долгоносиках паразитирует 11 видов браконид и 23 вида ихневмонов, всего 64 проц. найденных видов.
Из собранных нами 15 видов хищных мух из отряда Diptera определено всего 8 видов.
Среди собранных видов наибольшее значение в ограничении численности короедов имеют 3 вида из рода Medetera и 4 вида неопределенных нами более крупных мушек.
Из 15 видов жуков-хищников, собранных в подкоровых биоценозах, определено 13 видов.
За небольшим исключением, все насекомые паразиты и хищники выведены нами из личинок и куколок, собранных на ловчих модельных деревьях, а также на многочисленных обследованных стоящих и поваленных зараженных деревьях.
Биоценотические группировки скрытностволовых вредных насекомых с насекомыми хищниками и паразитами по данным анализа модельных деревьев
Всей серии выложенных нами в 1950 году в различных экологических условиях ловчих модельных деревьев свойственен несколько однообразный характер заселения вредителями. Наплыв короедов, по преимуществу раннего лета, помешал массовому заселению стволов усачами и другими вредителями. Только модели пихты и лиственницы заселились почти исключительно усачами и рогохвостами. На сосне наблюдалось несколько повышенное, против ели, заселение усачами (главным образом серым длинноусым) и рагием, а под тонкой корой — смолевкой Pissodes pini. На некоторых моделях ели в переходной высотной зоне усачи, за неимением свободного места на стволах, занятых типографом или чернобурым лубоедом, заселили в массе еловые сучья, находящиеся в затенении и поэтому не заселенные гравером. В числе этих усачей такие основные виды, заселяющие обычно стволы, как Tetropium castaneum, Semanotus undatus, Callidium aeneum, Saperda interrupta. Выведенные на ветвях жуки отличались значительно меньшими размерами, чем обычно.
Иной характер заселения отмечен на стоящих ослабленных деревьях. В первый же год они в массе заселялись усачами и лишь на следующий год к ним присоединялись короеды. Только на лиственнице усачи продолжают заселяться и в последующие годы до усыхания.
На поваленной лиственнице в верхней части ствола и на толстых сучьях поселяется иногда байкальский лесовик, но для поваленных сосен типично полное отсутствие заселения вредителями, тогда как на сучьях у стоящих сосен обычны и короеды рода Pityogenes, и смолевки Pissodes pini, P. piniphilus.
Поэтому поваленные ловчие модели сосны, ели и кедра представляли благодатный материал для изучения короедов и их врагов. Врагов усачей пришлось изучать преимущественно на деревьях этих пород, зараженных в стоящем положении и затем срубленных.
Серия модельных деревьев сосны взята нами в основных типах леса от предгорий до наиболее высоких точек района горной тайги, в участках относительно молодых гарей, старых гарей и негоревших насаждений.
Модели ели взяты от нижней границы ее распространения: (ручей Вторая Поперечная) в ельнике осочковом, до верхней границы: (Караульный камень) в сосняке-черничнике, а также в типе приручейной елово-пихтовой тайги. Две модели кедра взяты в пихтарнике-кисличнике зеленомошнике, одна из них на площади гари 1942 года, другая в негоревшем насаждении.
Ловчие модели лиственницы были выложены:
1) в предгорном районе (ручей Вторая Поперечная) в кв. 18 на старой гари в типе сосняка разнотравного, 2) в переходном поясе около старой западной границы заповедника в кв. 18, тоже на старой гари, 3) в районе горной тайги, на южном склоне от Второго Столба на гари 15-летней давности в сосняке-брусничнике.
В негоревшем насаждении взяты деревья, срубленные при устройстве квартальной сети в разных типах леса. Взята также стоящая зараженная модель лиственницы в негоревшем насаждении (сосняк-черничник около избушки «Музеянка» в кв. 48).
В качестве ловчих деревьев пихты использовался в достаточном количестве свежий ветровал, который отпиливался у шейки корня. Использовались и пихты, срубленные при устройстве квартальной сети. Взята одна стоящая модель пихты, зараженная усачами. И хотя по условиям заповедного режима 16 живых здоровых деревьев, разрешенных к вырубке, не позволили осуществить выкладку повторных циклов ловчих моделей в разные сроки, этот недочет был восполнен на ветровальных и стоящих зараженных деревьях.
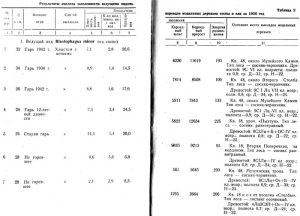
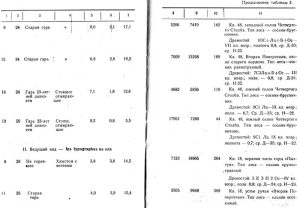
В таблице 2 приводятся данные анализа 16 моделей 1950 года. Сюда не вошли несколько моделей ели и кедра, так как накануне анализа были почти начисто окорены дятлами, поползнями и другими птицами. Результаты анализа ели, заселенно" Hylurgops glabratus, имеют относительное значение, так как чернобурый лубоед зимует в стадии личинок и нельзя вперед предсказать, какая часть их уцелеет до превращения в imago.
Пять первых моделей сосны, а также модели № 9 и № 15, как видно из таблицы 2, дают более или менее однообразные показатели заселения малым садовником; близкие к ним показатели имеет модель № 18, зараженная в стоячем положении. От них очень отличается модель № 19, тоже стоящая, но не усыхающая, еще свежая, и модели № 6 и № 7, выложенные в качестве ловчих в насаждениях, не тронутых пожарами. Плотность поселения короедов на первых семи моделях колеблется от 6,8 до 14,4 на 1 кв. дм коры, количество молодого поколения на 1 маточный ход от 1,4 до 3,4; продукция, то есть количество молодого поколения на 1 кв. дм, от 14,5 до 24,5; энергия размножения от 93 до 225 проц.
Средние показатели по типам леса для всех моделей сосны на гарях, а также для двух моделей в негоревших насаждениях сосняка-черничника приводятся в таблице 3.
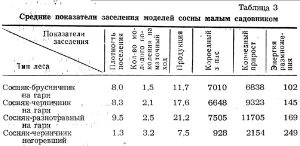
Как видно из таблицы 3, все показатели заселения находятся в тесной связи с типом леса, увеличиваясь по мере возрастания богатства почвы, а также в связи с бывшими пожарами. Только короедный запас в сосняке-брусничнике выделяется несколько преувеличенной цифрой за счет модели № 19, резко отличающейся от остальных моделей своим диаметром (55 см).
Анализ подтверждает наши наблюдения по биологии и экологии малого садовника. Этот лубоед, достаточно распространенный всюду, наиболее концентрируется в переходной полосе, а также вблизи водных потоков. Особенного размножения достигает на гарях. В нагоревших насаждениях плотность поселения в среднем на 1 кв.дм составляет 1,3 экз.; на гарях от 8 до 9,5. Короедный запас в негоревших насаждениях около 900 экз. на 1 дерево, а на гарях около 7000 экз.
Зато в негоревших насаждениях средняя цифра количества молодого поколения на 1 маточный ход — 3,2 экз., а на гарях от 1,5 до 2,5 экз. Естественно, что в негоревшем лесу, где запас короедов невелик, поселения их реже, пища для личинок обильнее, влажность воздуха выше, смягчаются колебания температуры, меньше естественных врагов короедов, и поэтому потомство малого садовника имеет более благоприятные условия для выживания, Поэтому и энергия размножения значительно выше в негоревших насаждениях. Все лежащие ловчие деревья сосны заселены лубоедом на протяжении 6 м от комля, а в области тонкой коры — смолевкой Pissodes pini, с плотностью поселения от 2 до 3 личинок на 1 кв.дм.
Самая высокая плотность поселения лубоеда отмечена на расстоянии 3 м от комля, а именно — в среднем для всех срубленных моделей на гарях 9,9 экз., несколько меньше на расстоянии 1 м — 9,5 экз., а всего меньше на расстоянии 5 м в области гонкой коры — 6,9 экз.
Толщина коры имеет большое значение для заселения короедами. В среднем по всем палеткам на 7 моделях плотность поселения составляет:
при толщине коры до 0,5 см-8,1
« » от 0,5 до 1,0 см - 8,8
« » от 1,0 до 1,5 см - 10,3
« » от 1,5 до 2,0 см - 9,2
« » от 2,0 до 2,5 см - 8,6.
Наибольшая плотность заселения наблюдается в среднем при толщине коры от 1 до 1,5 см. Эту особенность следует учитывать при выборе ловчих деревьев.
На лежащих ловчих моделях ели (см. таблицу 2) в нижнем горном поясе и переходной зоне поселяется типограф, а в горной тайге чернобурый лубоед и таежный лесовик. Плотность поселения типографа очень высокая, особенно на ветровальной ели, у которой часть корневой системы еще имеет связь с почвой. Вследствие перенаселенности количество молодых жуков на 1 маточный ход и энергия размножения снижается.

Модели ели, срубленные в предгорье (модель № 11) и в переходной зоне (модель № 8), почти не различаются по показателям плотности поселения, количеству молодого поколения на 1 маточный ход и продукции. На модели № 11, более старой по возрасту, комлевая часть до 9 м не заселена типографом, и поэтому здесь ниже короедный запас и короедный прирост, зато энергия размножения выше.
Отмечено, что на всех моделях ели короеды, в том числе и чернобурый лубоед, еще более чем на сосне, избегают селиться в области толстой трещиноватой коры. Поэтому ловчие деревья ели в расчете на короедов следует выбирать из более молодых по возрасту или же среди биотипов с гладкой корой.
Модель № 16 ели в негоревшем пихтарнике-кисличнике зеленомошнике и модель № 17 на гари в том же типе имеют очень высокую плотность поселения для этих видов. Но они не показательны в отношении потомства, проанализированного в стадии личинки.
Кроме вредителей, под корой живут и размножаются многие виды живущих за их счет насекомых хищников и паразитов, насекомых, питающихся остатками погибших личинок или детритом в ходах короедов; по-видимому, и личинки некоторых видов двукрылых питаются сочными частями луба.
В таблице 4 представлены данные по количеству на 1 кв.дм поверхности ствола молодого поколения всех видов насекомых, найденных под корой ловчих модельных деревьев, соответственно занимаемому ими положению в данной биоценотической группировке. Для лучшего понимания биоэкологических особенностей каждой категории насекомых таблица составлена с подразделением по секторам ствола. Секторы идут в порядке последовательного снижения степени освещенности, а вместе с тем и прогреваемости солнечной радиацией: верхний сектор; боковой, лучше освещаемый в течение дня; боковой, хуже освещаемый; нижний, обращенный к почве, наиболее затемненный.
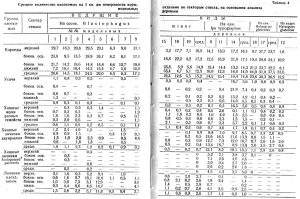
В биоценозах на моделях сосны наиболее многочисленен малый садовник; значительно ниже участие большого садовника и фиолетового лубоеда. В среднем из 11 моделей на 1 кв.дм. поверхности ствола насчитывается 15,2 экз. короедов. Личинок и куколок усачей в среднем всего 0,2 на 1 кв.дм, или 1 личинка приходится на 5 кв.дм коры.
Из врагов вредных насекомых и сожителей их на сосне наиболее многочисленны личинки наездников, в среднем 2,6 экз., и почти одинаковое количество жуков-хищников (imago и личинки); личинок хищных мух — 0,6 и 0,7 экз; хищников из отряда полужесткокрылых на сосне почти не отмечено.
На ели короеды также многочисленны; усачи хорошо заселили только модель № 14 — ветровальную. Наездников значительно меньше, чем в ходах малого садовника, всего 1,0 экз.; зато личинки хищных двукрылых достигают 70 проц. от числа короедов, а именно — 10,3 экз. Хорошо заметны на ели и хищные клопы, живущие за счет личинок короедов и мелких личинок усачей.
Биоценотические группировки в ходах Hylurgops glabratus на ели и кедре отличаются ничтожным количеством личинок наездников и двукрылых, жуков-хищников и хищных клопов.
Суммарное количество хищников и паразитов в ходах малого садовника составляет 26 проц. от числа короедов, в ходах типографа 109 проц., в ходах чернобурого лубоеда (личинки) всего 9 проц. Если даже ко времени окончательного развития в ходах чернобурого лубоеда останется половина наличного количества потомства, то и в этом случае их врагов будет не более 18 процентов.
По-видимому, паразиты и хищники могут существенно ограничивать потомство короедов только раннего лета, зимующих в стадии imago. Короеды позднего лета, например, Н. glabratus, приступают к размножению тогда, когда лет большинства паразитов и хищников заканчивается. Зато осенью от птиц страдает преимущественно потомство короедов, зимующих в стадии личинок.
На всех моделях сосны и ели, заселенных малым садовником и типографом, наблюдается антагонизм представителей различных отрядов хищных насекомых. Где под корой обильны личинки мух, там почти нет личинок наездников, и наоборот; часто сообщества тех и других имеют мозаичный характер, не смешиваясь друг с другом. Случаи истребления друг друга представителями различных отрядов насекомых хищников и паразитов нам почти не приходилось наблюдать. Преобладание в данном подкоровом биоценозе тех или иных групп насекомых, живущих за счет короедов, в основном зависит от экологической обстановки. Ксерофильные теплолюбивые виды поселяются обычно на стоящих деревьях. Гигрофилы предпочитают лежащие деревья, выбирая затененный сектор.
Анализ количественного распределения различных насекомых по секторам ствола приводит к следующему заключению. Заселенность лежащих сосен садовником зависит от суммы тепла и влаги, необходимых для его развития. Так, сильно затененные, с недостаточным количеством тепла модели в негоревших насаждениях или очень давно горевших заселяются слабо и более или менее равномерно; в среднем по трем верхним секторам имеем потомство в 7,3 экз. на 1 кв.дм, на нижнем секторе в среднем 4,9 экз.
На умеренно затененных соснах (модели № 1, № 4, № 5, № 18) оптимальные для развития жуков условия создаются на боковом освещенном секторе — 28,1 экз.; несколько хуже на верхнем, довольно хорошо освещенном секторе — 25,0 экз.; на боковом затененном секторе сказывается недостаток тепла — здесь 13,3 экз. и на нижнем, получающем мало тепла, всего 9,9 экз.
На равномерно и умеренно прогреваемых моделях достаточно приподнятых над почвой (модель № 2, № 3, № 9) довольно равномерное заселение — в среднем 16,6 экз.
Но на модели № 15 на прогалине в сосняке разнотравном, на полном солнечном освещении наилучшие условия для размножения создались в нижнем секторе — 33,4 экз.; несколько хуже на боковом затененном — 23,8 экз.; на боковом освещенном — 16,8 экз., а на верхнем секторе кора настолько пересохла, что большая часть потомства погибла в стадии личинки; достигли развития только 3,2 экз. на 1 кв.дм.
Как малый садовник на сосне, так и типограф на ели предпочитают достаточно хорошо, но не чрезмерно прогреваемые части ствола. На лучше освещенной модели № 8 на двух лучше прогреваемых секторах потомство типографа составляет 13,3 жуков на 1 кв. дм, на затененных 15,7 жуков. На моделях № 11 и № 14, находящихся в условиях худшей освещенности, имеем соответственно 18,5 и 13,5 жуков на 1 кв.дм. Даже такой типичный гигрофил, как чернобурый лубоед, на ели в сильно затененных холодных условиях лучше размножается на верхних секторах. На хорошо освещенной модели кедра он дал на верхнем секторе только 1,6 экз. на 1 кв.дм поверхности; на боковых секторах в среднем 9,5 экз. и на нижнем 33,6 экз.
Наиболее подвижная часть подкорового населения, а именно личинки и imago жуков-хищников в ходах малого садовника, распределяется аналогично наименее подвижным личинкам наездников; особенно насыщены ими затененные секторы, в частности затененный боковой.
В ходах типографа аналогичное явление обнаруживается только для малоподвижных личинок наездников. Хищных личинок Scoloposcelis pulchella больше на освещенных секторах; личинки двукрылых и жуки-хищники распределяются более или менее равномерно на освещенной и затененной частях ствола. Дело в том, что отдельные поселения малого садовника, включая маточные и личиночные ходы, даже при густом заселении, достаточно изолированы друг от друга и свободное передвижение хищников под корой затруднительно. В то же время поселения типографа соприкасаются весьма близко, да и кора ели быстрее разрушается и легче отстает от камбия, что и создает предпосылки к свободному передвижению под корой жуков-хищников. Личинки двукрылых по степени подвижности занимают среднее положение между личинками жу­ков и перепончатокрылых, они обладают способностью относительно свободно передвигаться во влажной среде между лубом и камбием в поисках очередной жертвы. Поэтому в их распределении по отдельным секторам ствола нет ясной закономерности ни на сосне, ни на ели.
Для анализа количественного соотношения насекомых на отдельных стволах взяты по две типичных модели сосны и ели (см. таблицу 5).
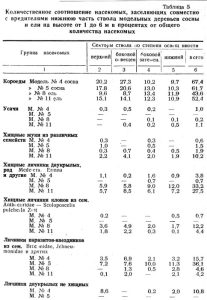
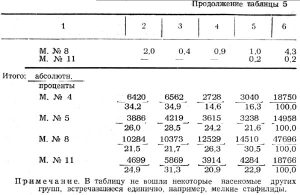
Модель № 4 сосны взята на старой гари в предгорном поясе в сосняке разнотравном; модель № 5 сосны на старой гари в переходной зоне тоже в сосняке разнотравном; модель № 8 ели в переходной зоне в ельнике крупнотравном; модель № 11 ели в предгорном поясе в ельнике осочковом; все четыре модели на склонах северо-восточной и восточной экспозиций.
Обе модели сосны заселены малым садовником на протяжении 6 м от комля. Модель № 4 имеет возраст 55 лет, диаметр на высоте груди — 24,5 см, высоту 22 м; модель № 5 в возрасте 80 лет имеет диаметр 25,5 см, высоту 21,3 м. Первая модель срублена на пологом склоне вблизи долины р.Лалетина и положена на низкие подкладки; вторая срублена вблизи долины ручья Второй Поперечной на крутом склоне; основная часть ее ствола вследствие изрезанности рельефа повисла над почвой на высоте до 1,2 м.
Как видно из таблицы 2, модель № 5 в предгорье заселилась короедами энергичнее, чем модель № 4 в переходной полосе. Плотность поселения на первой из них 14,4; на второй только 7,3.
Но к осени продукция короедов, короедный прирост и энергия размножения на модели № 5 оказалась, наоборот, ниже. Положение ствола модели № 5 способствовало более быстрому ее просыханию. В этих условиях создалась биоценотическая группировка ксерофильного характера, сходная с группировками на стоящем дереве. Личинки двукрылых, немногочисленные и на модели № 4 (4,5 проц.), здесь составили ничтожную величину: 0,7 проц. от общего количества насекомых, заселивших модель. Совершенно выпали из состава усачи и гигрофильная группа нехищных видов двукрылых, составляющая на модели № 4 — 10,8 проц. Хищные жуки, вообще немногочисленные в этом году, на модели № 5 представлены в большем количестве. Но паразиты-наездники достигли на модели № 5 36 проц. от общего числа насекомых.
Сопоставим на обеих моделях количество короедов и их врагов из трех групп хищников и паразитов (таблица 6).

Из этих данных видно, что на повалах сосны, заселенных малым садовником, группа хищных и паразитических насекомых составляет 24-38 проц., причем последняя цифра наивысшая для всех моделей сосны, короеды же составляют 76-62 проц.
В группе наездников мы не выделяли особо вторичных паразитов, исходя из того, что первичные выполнили свое дело и что количество вторичных паразитов примерно равно количеству первичных.
Абсолютное количество насекомых на модели № 11 ели значительно меньше, чем на модели № 8 (см. таблицу 5), так как типограф заселил первую лишь на протяжении от 9 м и выше. Но плотность поселения типографа почти одинакова на обеих моделях. Разница лишь в том, что на модели № 8 количество хищников и паразитов больше, что и привело к снижению количества короедов. Особенно много на модели № 8 личинок хищных мух и личинок Scoloposcelis pulchella.
Если исключить усачей и нехищных двукрылых, то на модели № 8 хищники и паразиты составляют вместо 54 проц., короеды же 46 проц.; на модели № 11 соответственно 47 проц. и 53 проц. Поэтому на модели № 8 энергия размножения типографа оказалась ниже, чем на модели № 11.
Процент наездников на обеих пробах почти одинаков, процент хищных жуков значительно выше на модели № 11; следовательно, на сокращение количества короедов на поваленных елях основное и решающее влияние оказывает группа насекомых хищных двукрылых, от численности которой зависит энергия размножения типографа.
В таблице 7 представлено среднее количество личинок мух на 1 кв.дм по отдельным биологическим группам видов.
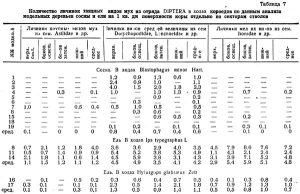
В первую группу выделены крупные белые личинки мух, видовой состав которых нами не определен. Лет мушек происходит в июне. Рост и питание их личинок наблюдались в 1950 году до конца первой декады сентября. Вначале они питаются личинками короедов, позже куколками и молодыми жуками. Особенно энергично уничтожают потомство короедов со времени достижения личин­кой мухи 1 см длины и до полного ее развития. В сентябре прячутся в ходах короедов, становятся неподвижными; к концу сентября коконируются в довольно крупном коричневом ложном коконе и зимуют под корой, частью зимуют без кокона. Личинки этих мух встречаются почти исключительно в ходах крупных видов короедов, главным образом типографа, реже садовников. Сравнительно немногочисленны под корой; держатся поодиночке.
Во вторую группу входят мушки величиной от 2,5 до 3,5 мм. Среди них наиболее многочисленны виды рода Medetera (семей­ство Dolychopodidae), из которых нами определены 4 вида (таблица 1). Самый распространенный вид М. signaticornis Lw. Род Medetera чрезвычайно многоядный. Найден нами на сосне, ели, кедре в ходах типографа, обоих садовников, двух полиграфов, гравера, нескольких видов Pityogenes и других. Чаще встречается все же на ели. Из сем. Lonchaeidae на сосне, иногда на ели, в ходах обоих садовников и типографа очень распространен Lonchaea seitneri Hend и реже Palloptera umbellatarum F. Личинки этих мух питаются личинками, иногда куколками короедов. Лет в июне. Развитие личинок Medetera происходит до сентября, затем они залегают между слоями волокон луба, согнув туловище пополам, и в таком положении зимуют. Пупарии образуются у одних видов с осени, у других весной. Вылет imago со второй половины июня до конца первой декады июля. Только imago Palloptera umbellatarum найдены были в период с 10 сентября по 30 октября в ходах малого садовника на сосне и в ходах гектографа на ели.
Личинки этих видов мушек держатся поодиночке, обладают способностью передвигаться между лубом и камбием и между лубяными волокнами независимо от ходов личинок короедов в поисках последних.
Личинки мушек из одного не определенного нами вида встречаются чаще сообществами по несколько экз., сообща используют убитую ими или другими хищниками личинку или жука. Эти личинки чрезвычайно подвижны и быстры. Не исключена возможность, что они имеют не одну генерацию в году, или же обладают очень растянутым летом, так как осенью мы находили в массе недокормившихся молодых личинок. Когда личинки крупных и средних мушек прекращают питаться, личинки вышеописанного не определенного нами вида питаются энергично до самых морозов, используя зимующих под корой короедов и их запоздавших личинок и куколок. Окукливаются без кокона.
В числе других хищных личинок под корой весьма обычны и личинки Erinna sincta De Geer.
Обычны под корой и личинки комариков из сем. Jtonidae и другие, не относящиеся к хищным, о чем свидетельствуют особенности строения головного конца личинок. Их сообщества часто встречались на незаселенных короедами деревьях; наряду с этим комарики охотно пользуются свежими ходами короедов и усачей для откладки яиц. Отмечено три вида этих комариков, не определенных нами; каждому виду свойственны свои биологические особенности. Зимуют в стадии личинок и imago; в пустотах под корой встречались значительные скопления их в стадии imago.
На моделях сосны личинок двукрылых мало (см. таблицу 7), на ели они весьма многочисленны, особенно в ходах типографа. Особенно хорошо представлены личинки средних и мелких видов мух, в среднем от 4,2 до 4,8 экз. на 1 кв.дм поверхности ствола. Личинки крупных мух располагаются главным образом на двух средне затененных секторах ствола; личинки средних без ясной закономерности, а личинки мелких располагаются на боковых и нижнем секторах ствола, проявляя некоторую гигрофильность. В ходах чернобурого лубоеда в связи с поздними сроками его лета (первая половина июля и даже позже) личинок мух мало, особенно крупных видов.
Под корой ели много личинок Scoloposcelis pulchella; заселяют они главным образом два лучше освещенных сектора (таблица 4).
Личинки наездников предпочитают хорошо освещенные стоящие деревья. Важнейшие, наиболее распространенные виды наездников, паразитирующие на скрытностволовых вредителях, многоядны; используют разнообразных хозяев и обладают широкой экологической пластичностью. Это — бракониды Atanycolus initiator Nees, паразитирующие на усачах, Coeloides melanotus Wesm, Coeloides abdominalis Zett на короедах; ихневмоны: Rhyssa persiasoria L, Ephialtes tuberculatus Fourc — на усачах, рогохвостах, долгоносиках. Каждому из этих наездников присущи свои особенности в выборе хозяев.
Бракониды Atanycolus являются эктопаразитами усачей, выбирают хозяев из триб Asemini и Callidini, а именно: Tetropium castaneum, Т. gracilicorne, Semanotus undatus, Callidium aencum, C. violaceum и других. Их желтоватые полупрозрачные овально-продолговатые коконы легко отличить по плоской верхней и нижней поверхности и окружающим швам; расположены в ходах усачей, всегда только под корой.
Браконид Coeloides melanotus всюду многочисленен в ходах малого садовника, шестизубого короеда, типографа, фиолетового лубоеда, пушистого полиграфа, гектографа и др.
Ихневмон Rhyssa persuasoria выбирает хозяев, личинки которых уходят глубоко в древесину, в том числе — Monochamus, Sirex, Paururus. Его коконы располагаются обычно глубоко в древесине в ходах проточенных хозяином.
Ихневмон Ephialtes tuberculatus, наоборот, располагает свои удлиненные светлые коконы под корой. Хозяева его — все виды Monochamus, Acanthocinus aedilis, Rhagium inquisitor, R. mordax, Tetropium castaneum, Hylobius abietis, не считая многих других хозяев, не охваченных нашими исследованиями.
Среди усачей наиболее поражаются наездниками Tetropium catsaneum (9 видов) Semanotus undatus (8 видов), Monochamus urussovi (7 видов) и Т. gracilicorne (6 видов).
На разных стадиях развития личинки отдельных видов усачей заражаются разными видами наездников, смотря по размерам последних и их биологическим особенностям. Мелкие виды наездников выбирают более мелких молодых личинок усачей, а более крупные — крупных.
При наличии в данном месте ствола личинок усачей разного возраста, например, личинок Tetropium gracilicorne на лиственнице, личинки из ранее отложенных яичек заражаются Ephialtes tuberculatus, личинки же из потомства запоздавших особей этого усача заражаются Itoplectis maculator и другими. Личинки садовников и типографа заражаются хальцидами и мелкими видами браконид в достаточно развитом состоянии, перед окукливанием. По-видимому, не всегда личинка браконида, являющаяся наружным паразитом, может использовать нескольких личинок короеда. А так как личинка последнего, после присасывания паразита, становится неподвижной и перестает питаться, то она должна быть достаточно крупной, чтобы обеспечить развитие паразита. Личинка паразита развивается быстро, но, закоконировавшись, она очень длительный срок (около 8-9 месяцев) находится в коконе до своего окукливания.
Из 8 найденных нами видов сем. Chalcididae — 7 видов широко распространены на короедах, но определить их нам не удалось, поскольку специалист-систематик, которому экземпляры их были высланы на определение, не смог найти для этого времени.
Большинство наездников, в особенности бракониды, обладают ограниченным во времени летом. Лет некоторых видов растянут в течение всего сезона. Пример — Rhyssa persuasoria, начинающая лет в первых числах июня и заканчивающая его в конце сентября. Несомненно, это является результатом длительного приспособления паразитов к генерации выбираемых ими хозяев.
В 1950 году у большинства мелких видов наездников лет начался 20 июня, когда у короедов раннего лета (садовники, фиолетовый лубоед и др.) личинки достигли уже достаточной степени развития. К этому же сроку относится и массовый лет Rhyssa persuasoria и других крупных наездников. Массовый лет продолжался месяц. Вероятно, этим объясняется сравнительно невысокая зараженность короедов позднего лета, как например, H. glabratus.
В лабораторных условиях продолжительность жизни самок всех наездников установлена в 1 месяц; самцы живут значительно меньше. В лаборатории же установлено, что из серии собранных коконов любого наездника, если имеются самцы, они вылетают первые и лишь спустя некоторое время, иногда 2-3-5 дней начинается вылет самок. То же явление наблюдалось и в природных условиях при массовом лете. Над некоторыми поваленными зараженными деревьями, кружатся роями одни самцы. Лет самцов продолжается недолго, и в конце июня, начале июля встречаются уже почти исключительно самки.
Никакой почти связи с цветущими растениями у наездников-паразитов скрытностволовых вредителей нами не установлено. Они встречались нам или на лету, или на стволах деревьев, заселенных вредителями.
В жаркое время дня летают только единичные крупные наездники Rhyssa и Ephialtes и откладывают яички. Большинство паразитов, и, прежде всего, мелкие бракониды и хальциды, начинают свой лет с 10 ч. утра и к 11-12 часам кончают. Вечерний лет происходит с 5-6 ч. вечера до вечерней росы. Усиленный лет наблюдается перед дождями и грозами, когда «парит». По нашим наблюдениям оптимум их лета при 18-22° С и относительной влажности воздуха 60-70 проц.
Вылет молодого поколения наездников в лабораторных условиях происходил при температуре не ниже +10°, но вяло; оптимальная же температура около 22° С при достаточно высокой влажности воздуха.
Этими биологическими свойствами наездники отличаются от двукрылых. Многие виды мух интенсивно летают над зараженными деревьями и откладывают яички в самые жаркие дни июля, когда относительная влажность воздуха падает до 40 проц. и ниже.
Роль жуков-хищников в истреблении короедов весьма значительна. Хорошо известный истребитель короедов Cleroides formicarius уничтожает imago короедов в момент их втачивания в кору для откладки яиц. Его розовая личинка истребляет большое количество личинок и куколок садовников, прогрызая в коре самостоятельные ходы в погоне за личинками короедов, когда последние углубляются в толщу коры для окукливания, как это имеет место в случае очень толстой коры.
Особое значение имеют жуки яйцееды, забирающиеся в маточные ходы садовников, типографа, гравера, и др. и выедающие там отложенные яички. В заповеднике к ним относятся Cilistosoma oblongum, Laemophloeus abietis, Rhizophagus dispar, R. depressus, Hypophloeus suturalis, H. linearis и другие.
По нашим наблюдениям 1949 года на стоящей зараженной модели ели в урочище «Слизнево» на гари 1945 года в гнездах гравера в массе находились жуки Rhizophagus depressus. Потомство же гравера погибло почти полностью, частью в стадии яйца, частью в стадии молодых личинок, истребленных личинками хищника.
Очень большое значение имеют и не определенные нами мелкие виды стафилид. Их личинки истребляли множество молодых личинок различных короедов, предоставляя остатки своей пищи личинкам других видов насекомых. Случалось видеть, как личинка стафилиды уничтожала и хищных личинок двукрылых.
Нередки в ходах короедов и личинки верблюдки Rhaphidia sp.
В области крон стоящих усыхающих деревьев, где гнездятся мелкие виды короедов рода Pityogenes и Pityophthorus, в их ходах также размножается много врагов, в достаточной степени ксерофильных.
Из ветвей или, заселенных гравером, нами были выведены мелкие виды хальцид в количестве до 60 проц. от числа молодых жуков гравера. На сосновых ветвях, заселенных Pityogenes irkutensis. P. bidentatus, такого обилия хальцид не отмечалось, самое большее — 24 проц. от числа выведенных короедов. Обычно же на еловых ветвях в ходах Pityogenes chalcographus, P. saalasi и других отмечалось 10-15 проц., хальцид, на сосновых — 5-7 проц. На ветвях ели найдены и мелкие бракониды: Sphathium rubidus Ross и Ecphylus hylesini Ratz.
Из двукрылых на ветвях хвойных выведены в относительно небольшом количестве мушки из сем. Dolychopodidae и других.
Значительно выше численность хищных жуков, главным образом Rhizophagus depressus F и Hypophloeus lineare F. Нередко к ходам гравера примыкает на ветвях и короед Crypturgus cinereus, являющийся, как известно, по отношению к другим короедам хищником, поедающим их яйца.
Проведенные нами исследования биологических и экологических свойств насекомых — врагов скрытностволовых вредителей — еще очень недостаточны, чтобы дать хозяйственные указания по биологическому методу борьбы с вредителями. Особенно нуждаются в изучении и уточнении видового состава хальциды, из которых некоторые виды являются вторичными паразитами.
Среди исследованных нами насекомых не установлены виды быстро размножающиеся. Для всех типична одногодичная генерация.
Закоконировавшиеся личинки паразитов значительно лучше переносят все неблагоприятные условия, чем личинки вредителей; хорошо выдерживают морозы, избыточную влажность и, наоборот, длительную сухость даже при отсутствии кокона. .
Поэтому в целях борьбы со скрытностволовыми вредителями биологическим методом, не касаясь вопроса размножения наездников в лабораторных условиях, не представляет трудностей создавать запасы личинок паразитов, собирая коконы при проведении осенью санитарных рубок. Использование их на свежих гарях вполне возможно, так как собранные осенью коконы прекрасно сохраняются в кусках коры при зимнем хранении в прохладном помещении, но желательно при температуре не ниже −10° С. При хранении в комнатной температуре от 10° до 20° С вылет насекомых происходит в январе-марте месяце.
Можно также собирать куски коры с коконами непосредственно перед окукливанием паразитов или в период окукливания в мае месяце и переносить их на площадь гари.
Перед сбором коконов мелких браконид необходимо их внимательно просматривать, так как сбор коконов, зараженных вторичными паразитами, бесполезен. Такие коконы легко отличить по мелким и расположенным без всякого порядка отверстиям, проделанным в них вылетевшими сверхпаразитами. В отличие от первичных паразитов последние обладают укороченным сроком генерации и выводятся летом.
Личинки хищных мух при снятии коры и нарушении свойственных им естественных условий, как правило, гибнут. Однако хранение их в ложно-коконах при определенных условиях влажности и температуры вполне возможно.
В заключение следует отметить, что вопросы биологического метода борьбы со скрытностволовыми вредителями нуждаются не только в специальных экспериментах в природе, но и в наличии соответственно оборудованной лаборатории.
Literature
1. Воронцов А.И. Использование хищных жуков для борьбы с короедами. Результаты работ ВНИИЛХ за 1941-1945 гг., вып. 27, М.-Л., 1949.
2. Гречкин В.П. Биологический метод борьбы с вторичными вредителями леса. Результаты работ ВНИИЛХ за 1941-1945 гг., вып. 27. М.-Л., 1949.
3. Гусев В.И. Полезные насекомые, встречающиеся на деревьях, заселенных короедами. Изв. Ленинградского лесного ин-та, XXXYII, 1928.
4. Мейер Н.Ф. Биологический метод борьбы с вредными насекомыми. Труды 3-го Всероссийского съезда энтомологов и фитопатологов, М., 1921.
5. Мейер Н.Ф. Об учете значения паразитических перепончатокрылых в динамике вредных насекомых. Известия Гос. Ин-та опытной аграномии, т.YII, № 3-4, М., 1929.
6. Мейер Н.Ф. Определитель паразитических перепончатокрылых СССР и сопредельных стран. Вып I — 1933, в. II — 1933, в. III — 1934, в. IY — 1935, в. Y и YI — 1936 г., М.-Л.
7. Никитюк А.И. Хищные и паразитические насекомые, как регуляторы вредоносной деятельности и распространения короедов хвойного леса. Бюллетень МОИП, отд. биологии, т.LYII (5), 1952.
8. Померанцев Д. Биологические заметки о жуках, полезных в лесоводстве, живущих под корой деревьев. Р. Энт. Обозр., т. II — 1902 г., т. III — 1903 г., т. IY — 1904 г., СПБ.
9. Померанцев Д. К познанию вредных и полезных насекомых, водящихся на ели (Picea excelsa) Изв. СПБ Лесного института, в. YIII СПБ. 1902.
10. Попов В.В. Перепончатокрылые — Hymenoptera. «Животный мир СССР», т. 4. Изд. АН СССР, М.-Л., 1953.
11. Старк Н.Н., Старк В.Н., Белановский И.Д. Методы и техника борьбы с вредными лесными насекомыми, М., 1932.
12. Теленга Н.А. Фауна СССР. Насекомые перепончатокрылые, Y, 2. Braconidae. Изд. АН СССР. М.-Л., 1936,
13. Теленга Н.А. Фауна СССР. Насекомые перепончатокрылые, Y. 3. Bracoridae. Изд. АН СССР. М.-Л., 1941.
14. Шевырев И. Паразиты и сверхпаразиты из мира насекомых. СПБ, 1912.
15. Штакельберг А.А. Определитель мух Европейской части СССР. Изд. зоологич. ин-та АН СССР, 7, М.-Л., 1933.
16. Штакельберг А.А. Двукрылые — Diptera. «Животный мир СССР», т. 4. Изд. АН СССР, М.-Л., 1953.
Owner →
Offered →
Collection →
Севастьянова Татьяна Петровна
Севастьянова Татьяна Петровна
Труды государственного заповедника